

Buy feldene online from canada
Finally arthritis in fingers pain relief order feldene with a visa, the blob pathway receives input from the subset of ganglion cells that are neither M-type cells nor P-type cells. Many neurons in the blobs are color selective, so they might be involved in the analysis of object color. The description earlier is the simple version of the story; the reality is more complicated. Research has shown that the three proposed pathways do not keep magnocellular, parvocellular, and koniocellular signals separate; instead they mix. Also, receptive field properties such as orientation and color tuning are found across the proposed pathways. Thus, it is not the case that magnocellular, parvo-interblob, and blob neurons are strictly segregated and have entirely unique receptive field properties. Collectively, these observations are consistent with the notion that this is an output pathway particularly involved in navigation and the analysis of motion. We will see later that beyond the striate cortex, there appear to be two major pathways handling different types of visual information, one extending toward the parietal lobe that deals with motion and the other involved with color and form extending toward the temporal lobe. Receptive fields in primary visual cortex range from a fraction of a degree to several degrees across, and nearby cells have receptive fields that overlap a great deal. For these reasons, even a small spot of light will activate thousands of V1 neurons. Hubel and Wiesel showed that the image of a point in the visual field falls within the receptive fields of neurons in a 2 2 mm chunk of macaque striate cortex. Each cortical module contains ocular dominance columns, orientation columns, and cytochrome oxidase blobs to fully analyze a portion of the visual field. The idealized cube shown here differs from the actual arrangement, which is not as regular or orderly. Ocular dominance columns Thus, Hubel and Wiesel argued that a 2 2 mm chunk of striate cortex is both necessary and sufficient to analyze the image of a point in space: necessary because its removal would leave a blind spot for this point in the visual field, and sufficient because it contains all the neural machinery required to analyze the form and color of objects viewed through either eye. Because of the finite size of receptive fields and some scatter in their locations, a cortical module processes information about a small patch of the visual field. We can think of a visual scene being simultaneously processed by these modules, each "looking" at a portion of the scene. Beyond V1 lie another two dozen distinct extrastriate areas of cortex that have unique receptive field properties. The contributions to vision of these extrastriate areas are still being vigorously debated. The dorsal stream appears to serve the analysis of visual motion and the visual control of action. The ventral stream is thought to be involved in the perception of the visual world and the recognition of objects. These processing streams have primarily been studied in the macaque monkey brain, where recordings from single neurons can be made. Early visual areas including V1, V2, V3, V3A, and V4 are retinotopically organized. Higher temporal lobe areas involved in face and object recognition are not retinotopic. However, each extrastriate stream receives some amount of input from all the pathways in the primary visual cortex. The Dorsal Stream the cortical areas composing the dorsal stream are not arranged in a strict serial hierarchy, but there does appear to be a progression of areas in which more complex or specialized visual representations develop. Projections from V1 extend to areas designated V2 and V3, but we will skip farther ahead in the dorsal stream. This cortical area is arranged into direction-of-motion columns analogous to the orientation columns in V1. For example, if electrical stimulation is applied to cells in a direction column preferring rightward movement, the monkey makes behavioral decisions suggesting that it has perceived motion in that direction. Navigation: As we move through our environment, objects stream past our eyes, and the direction and speed of objects in our peripheral vision provide valuable information that can be used for navigation. Directing eye movements: Our ability to sense and analyze motion must also be used when we follow objects with our eyes and when we quickly move our eyes to objects in our peripheral vision that catch our attention. Motion perception: We live in a world filled with motion, and survival sometimes depends on our interpretation of moving objects. The clearest case was reported in 1983 by Josef Zihl and his colleagues at the Max Planck Institute for Psychiatry in Munich, Germany. Although some ill effects of the stroke were evident, such as difficulty naming objects, neuropsychological testing showed the patient to be generally normal and to have relatively normal vision, except for one serious deficit: She appeared to be incapable of visually perceiving motion. Before you decide that not seeing motion would be a minor impairment, imagine what it would be like to see the world in snapshots. More ominously, she had trouble crossing the street-one moment she would perceive cars to be in the distance, and the next moment they would be right next to her. The implication of this case is that motion perception may be based on specialized mechanisms located beyond the striate cortex in the dorsal stream. V4 receives input from the blob and interblob regions of the striate cortex via a relay in V2. Neurons in area V4 have larger receptive fields than cells in the striate cortex, and many of the cells are both orientation selective and color selective. Although there is a good deal of ongoing research into the function of V4, this area appears to be important for both shape perception and color perception. If this area is damaged in monkeys, perceptual deficits involving both shape and color result. A rare clinical syndrome in humans known as achromatopsia is characterized by a partial or complete loss of color vision despite the presence of normal functional cones in the retina. People with this condition describe their world as drab, consisting of only shades of gray. Collaborating with an undergraduate, Josh McDermott, and post-doc, Marvin Chun, I spent some of the happiest moments of my life lying inside the scanner bore, biting on a bite bar, and watching Marvin and Josh (upside down) through the mirror over my forehead, as they operated the scanner from out in the console room. What astonishing good fortune to get to use this amazing machine to explore the largely uncharted territory of human visual cortex! We started out by trying to find brain regions engaged in the perception of object shape. As I did not have a grant to pay for scan time, I knew that my scanning privileges would not continue unless I hit a home run-and quickly. The extensive behavioral literature on normal and braindamaged individuals strongly suggested that a special part of the brain might exist for face perception. What they had not investigated was whether this response was specific to faces, or whether the same region might also be engaged during perception of other complex visual stimuli. This specificity question connected directly to one of the most long-standing and fierce debates in the histories of both cognitive science and neuroscience: To what extent are the mind and brain composed of special-purpose mechanisms, each processing a specific kind of information We figured that if a special-purpose part of the brain existed that was selectively involved in face perception, it should produce a stronger response when people look at faces than when they look at objects. We then scanned subjects while they looked at these face photos and at photos of common objects. To our delight, we found that in almost everyone the image showed a nice clear blob on the lateral side of the fusiform gyrus, primarily in the right hemisphere, where the statistics told us that the response was higher when people were looking at faces than when they were looking at objects. To deal with this anatomical variability, plus to make our statistical analyses bulletproof, we split the data for each subject in half, using half the data to find the region with our faces-versusobjects contrast, and the other half to quantify the response with the coexistence of color-sensitive and shape-sensitive cells in the ventral stream, achromatopsia is usually accompanied by deficits in form perception. Some researchers have proposed that V4 is a particularly critical area for color and form perception, but the lesions associated with achromatopsia are generally not limited to V4, and severe visual deficits appear to require damage to other cortical areas in addition to V4. Beyond V4 in the ventral stream are cortical areas that contain neurons with complicated spatial receptive fields. One reason this area is of particular interest is that it appears to be the farthest extent of visual processing in the ventral stream.
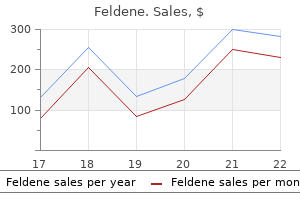
Order generic feldene line
To describe your location in the city arthritis knee rest cheap feldene online, you would use points of reference such as north, south, east, and west and up and down. The same is true for the brain, except that the terms-called anatomical references-are different. We begin with the rat because it is a simplified version that has all the general features of mammalian nervous system organization. In the head lies the brain, and the spinal cord runs down inside the backbone toward the tail. The direction pointing up is known as dorsal (from the Latin for "back"), and the direction pointing down is ventral (from the Latin for "belly"). Despite differences in complexity, the brains of all these species have many features in common. The brains have been drawn to appear approximately the same size; their relative sizes are shown in the inset on the left. Caud al Rost ral (a) Midsagittal Thus, the rat spinal cord runs anterior to posterior. The top side of the spinal cord is the dorsal side, and the bottom side is the ventral side. With just a few exceptions, most structures within the nervous system come in pairs, one on the right side and the other on the left. The invisible line running down the middle of the nervous system is called the midline, and this gives us another way to describe anatomical references. Structures closer to the midline are medial; structures farther away from the midline are lateral. In other words, the nose is medial to the eyes, the eyes are medial to the ears, and so on. In addition, two structures that are on the same side are said to be ipsilateral to each other; for example, the right ear is ipsilateral to the right eye. If the structures are on opposite sides of the midline, they are said to be contralateral to each other; the right ear is contralateral to the left ear. To view the internal structure of the brain, it is usually necessary to slice it up. In the language of anatomists, a slice is called a section; to slice is to section. Although one could imagine an infinite number of ways we might cut into the brain, the standard approach is to make cuts parallel to one of the three anatomical planes of section. The two other anatomical planes are perpendicular to the sagittal plane and to one another. A single section in this plane could pass through both eyes or both ears but not through all four at the same time. Notice that it is clearly split down the middle into two cerebral hemispheres, separated by the deep sagittal fissure. In general, the right cerebral hemisphere receives sensations from, and controls movements of, the left side of the body. Similarly, the left cerebral hemisphere is concerned with sensations and movements on the right side of the body. Lying behind the cerebrum is the cerebellum (the word is derived from the Latin for "little brain"). While the cerebellum is in fact dwarfed by the large cerebrum, it actually contains as many neurons as both cerebral hemispheres combined. The cerebellum is primarily a movement control center that has extensive connections with the cerebrum and the spinal cord. In contrast to the cerebral hemispheres, the left side of the cerebellum is concerned with movements of the left side of the body, and the right side of the cerebellum is concerned with movements of the right side. The brain stem forms the stalk from which the cerebral hemispheres and the cerebellum sprout. The brain stem is a complex nexus of fibers and cells that in part serves to relay information from the cerebrum to the spinal cord and cerebellum, and vice versa. However, the brain stem is also the site where vital functions are regulated, such as breathing, consciousness, and the control of body temperature. Indeed, while the brain stem is considered the most primitive part of the mammalian brain, it is also the most important to life. One can survive damage to the cerebrum and cerebellum, but damage to the brain stem is usually fatal. The spinal cord is encased in the bony vertebral column and is attached to the brain stem. Axons enter and exit the spinal cord via the dorsal and ventral roots, respectively. A transection of the spinal cord results in anesthesia (lack of feeling) in the skin and paralysis of the muscles in parts of the body caudal to the cut. Paralysis in this case does not mean that the muscles cannot function, but they cannot be controlled by the brain. The spinal cord communicates with the body via the spinal nerves, which are part of the peripheral nervous system (discussed below). Spinal nerves exit the spinal cord through notches between each vertebra of the vertebral column. Charles Bell showed that the ventral root contains axons carrying information away from the spinal cord-for example, to the muscles that jerk your foot away in response to the pain of the thumbtack. The somatic motor axons, which command muscle contraction, derive from motor neurons in the ventral spinal cord. The somatic sensory axons, which innervate and collect information from the skin, muscles, and joints, enter the spinal cord via the dorsal roots. Visceral motor fibers command the contraction and relaxation of muscles that form the walls of the intestines and the blood vessels (called smooth muscles), the rate of cardiac muscle contraction, and the secretory function of various glands. Derived from the Latin, afferent ("carry to") and efferent ("carry from") indicate whether the axons are transporting information toward or away from a particular point. The Cranial Nerves In addition to the nerves that arise from the spinal cord and innervate the body, there are 12 pairs of cranial nerves that arise from the brain stem and innervate (mostly) the head. Each cranial nerve has a name and a number associated with it (originally numbered by Galen, about 1800 years ago, from anterior to posterior). Many cranial nerves contain a complex mixture of axons that perform different functions. The cranial nerves and their various functions are summarized in the chapter appendix. It is protected by three membranes collectively called the meninges (singular: meninx), from the Greek for "covering. While there normally is no space between the dura and the arachnoid, if the blood vessels passing through the dura are ruptured, blood can collect here and form what is called a subdural hematoma. Choroid plexus Rostral Subarachnoid space the pia mater, the "gentle mother," is a thin membrane that adheres closely to the surface of the brain. Along the pia run many blood vessels that ultimately dive into the substance of the underlying brain. The fluid-filled caverns and canals inside the brain constitute the ventricular system. As we will see, understanding the organization of the ventricular system holds the key to understanding how the mammalian brain is organized. New Views of the Brain For centuries, anatomists have investigated the internal structure of the brain by removing it from the skull, sectioning it in various planes, staining the sections, and examining the stained sections. However, because the skull is soft and not completely formed, the head will expand in response to the increased intracranial fluid, sparing the brain from damage. Often this condition goes unnoticed until the size of the head reaches enormous proportions. In adults, hydrocephalus is a much more serious situation because the skull cannot expand, and intracranial pressure increases as a result. The soft brain tissue is then compressed, impairing function and leading to death if left untreated. Typically, this "obstructive" hydrocephalus is also accompanied by severe headache, caused by the distention of nerve endings in the meninges. The trick is to soak the brain in a solution that replaces light-absorbing lipids with a water-soluble gel that turns the brain transparent. This, to say the least, limits the usefulness of such anatomical methods for diagnosing neurological disorders in living individuals.
Diseases
- Benign familial infantile epilepsy
- Poikilodermia alopecia retrognathism cleft palate
- Deafness hyperuricemia neurologic ataxia
- Scleromyxedema
- Chronic polyradiculoneuritis
- Circumscribed cutaneous aplasia of the vertex
Order feldene 20 mg on-line
Although affected children appear to be normal at birth arthritis relief cream australia purchase discount feldene online, symptoms gradually appear over the course of the first 3 years. Among the signs first noticed by the parents of autistic children are a failure to speak by 16 months of age, poor eye contact, an inability to play with toys, an obsessive attachment to a toy or object, and a failure to smile. Although all individuals with an autism diagnosis will show these traits, the severity varies considerably from one person to the next, as does the association or "comorbidity" with other diagnosable disorders such as intellectual disability and seizures. Individuals at one end of the spectrum may never develop language and exhibit severe cognitive impairment. At the other end, individuals may grow up to be socially awkward but intellectually gifted. In some cases, the gene mutations conferring risk for autism occur de novo, meaning that they occur sporadically either in the sperm or egg cells of the parents. One risk factor for such sporadic mutations is advanced parental age, especially of fathers. The diversity of genetic etiologies partly explains why the symptoms vary so much from one person to the next. For example, researchers recently discovered in postmortem brain samples from autistic children that small patches of frontal cortex had disorganized cortical layers which, as we have learned in this chapter, are formed early in development. Imaging studies have shown that autistic children also tend to have accelerated growth of the brain, both gray and white matter, after birth. This finding suggests the brains of autistic infants have too many neurons and too many axons, although changes in glia are also possible. Brain growth is controlled by balancing the genesis and destruction of cells, axons, and synapses and the proteins that comprise them. Mutations that bring this process out of balance, by excessive genesis or reduced destruction, could lead to the abnormal brain growth that is ultimately expressed as the impairments in behavior, communication, and social interactions that characterize autism. Neuroscientists hope that understanding how the brain normally becomes wired together will suggest therapies to correct the altered trajectory of brain growth in children at risk for autism. By knocking this gene out in mice and fruit flies, researchers have been able to identify how brains function differently with this mutation. These studies have raised the tantalizing possibility that the veil of autism and intellectual disability might be lifted in some cases with appropriate drug therapy. The development of proper brain function requires a careful balance between the genesis and elimination of cells and synapses (Box 23. Cell Death Entire populations of neurons are eliminated during pathway formation by a process known as programmed cell death. After axons have reached their targets and synapse formation has begun, there is a progressive decline in the number of presynaptic axons and neurons. Cell death reflects competition for trophic factors, life-sustaining substances that are provided in limited quantities by the target cells. The input neurons are believed to compete with one another for limited quantities of trophic factors produced by the target neurons. Most of the receptors are neurotrophin-activated protein kinases, called trk receptors, that phosphorylate tyrosine residues on their substrate proteins (recall phosphorylation from Chapter 6). The description of cell death during development as "programmed" reflects the fact that it is actually a consequence of genetic instructions to self-destruct. The important discovery of cell death genes by Robert Horvitz at the Massachusetts Institute of Technology was recognized with the 2004 Nobel Prize. It is now understood that neurotrophins save neurons by switching off this genetic program. The expression of cell death genes causes neurons to die by a process called apoptosis, the systematic disassembly of the neuron. Apoptosis differs from necrosis, which is the accidental cell death resulting from injury to cells. Changes in Synaptic Capacity Each neuron can receive on its dendrites and soma a finite number of synapses. Throughout the nervous system, synaptic capacity peaks early in development and then declines as the neurons mature. For example, in the striate cortex of all species examined so far, the synaptic capacity of immature neurons exceeds that of adult cells by about 50%. In other words, visual cortical neurons in the infant brain receive one-and-a-half times as many synapses as do the neurons in adults. Yale University scientists Jean-Pierre Bourgeois and Pasko Rakic conducted a detailed study to address this question in the striate cortex of the macaque monkey. They discovered that synaptic capacity was remarkably constant in the striate cortex from infancy until the time of puberty. However, during the subsequent adolescent period, synaptic capacity declined sharply-by almost 50% in just over 2 years. A quick calculation revealed the following startling fact: the loss of synapses in the primary visual cortex during adolescence occurs at an average rate of 5000 per second. Simply blocking a subset of receptors with -bungarotoxin can also stimulate synapse elimination. Silencing the activity of the muscle fiber leads to a retention of polyneuronal innervation, while stimulation of the muscle accelerates the elimination of all but one input. The answer appears to be insufficient receptor activation in an otherwise active muscle. Another arrangement is that neuron A provides one synapse and neuron B provides five. The target cell receives the same number of synapses in both cases, but the innervation pattern has changed. There is abundant evidence for widespread synaptic rearrangement in the immature brain. Unlike most of the earlier steps of pathway formation, synaptic rearrangement occurs as a consequence of neural activity and synaptic transmission. In the visual system, some of this activity-dependent shaping of connections occurs prior to birth in response to spontaneous neuronal discharges. However, significant activity-dependent development occurs after birth and is influenced profoundly by sensory experience during childhood. Thus, we will find that the ultimate performance of the adult visual system is determined to a significant extent by the quality of the visual environment during the early postnatal period. In a very real sense, we learn to see during a critical period of postnatal development. Macaque monkeys and cats were used by Hubel and Wiesel as models for studies of activity-dependent visual system development because, like humans, both of these species have good binocular vision. Recent studies have used rodents because they are better suited for investigation of the underlying molecular mechanisms. Synaptic Segregation the precision of wiring achieved by chemical attractants and repellents can be impressive. In some circuits, however, the final refinement of synaptic connections appears to require neural activity. Somewhat later, the ipsilateral projection arrives and intermingles with the axons of the contralateral eye. Then the axons from the two eyes segregate into the eye-specific domains that are characteristic of the adult nucleus. Since segregation occurs in the womb, prior to the development of photoreceptors, the activity cannot be driven by light stimulation. Studies by Carla Shatz and her colleagues at Stanford University indicate that ganglion cells fire in quasisynchronous "waves" that spread across the retina. The origin of the wave and its direction of propagation may be random, but during each wave, the activity in a ganglion cell is highly correlated with the activity of its nearest neighbors. And because these waves are generated independently in the two retinas, the activity patterns arising in the two eyes are not correlated with respect to each other. This hypothetical mechanism of synaptic plasticity was first articulated by Canadian psychologist Donald Hebb in the 1940s. Consequently, synapses that can be modified in this way are called Hebb synapses, and synaptic rearrangements of this sort are called Hebbian modifications.
Buy discount feldene 20 mg on line
That being said arthritis neck facet disease buy 20mg feldene free shipping, we must also account for the inherent toxicity of the nanoparticles themselves when developing nanoparticle-based therapies. A balance must be struck when deciding appropriate nanoparticle dosage to allow for maximum effect with the least chance of side effects. To encapsulate things, the use of nanoparticle-based therapies is an exciting new field in wound treatment, with endless prospects and possibilities. Whether these therapies involve the delivery of substances or promote wound healing due to the inherent properties of the nanoparticles, there are promises of achieving greater efficacy and specificity, with fewer systemic side effects. In addition, compared to conventional antibiotics, nanoparticle-based antibacterial therapies may also be less likely to result in bacteria developing resistance. Current treatments that involve nanoparticles have also demonstrated successful track records. However, much is yet to be discovered regarding the effects of nanoparticles on the human body, and for this reason, the development of nanoparticle-based therapies should be undertaken with a reasonable amount of caution. Working to improving the efficacy of nanoparticle wound treatments should go hand-in-hand with investigating the long- and short-term effects of said treatments, as well as the mechanisms underlying them. That being said, we are currently approaching what can be considered the golden age of nanotechnology, with research occurring at an exponential rate. With new ways of manipulating nanomaterials into nanoparticles emerging rapidly, it seems reasonable to be optimistic about developing methods of synthesizing nanoparticles, whether in vitro or in situ, that can maximize efficacy without compromising the safety of the patient, the people administering the treatment, or the environment in general. Further development in this emerging field will definitely have a positive impact on the treatment of wounds, especially chronic wounds, which pose a significant burden on quality of life and the healthcare industry. Lipid-polymer hybrid nanoparticles as a new generation therapeutic delivery platform: a review. Plateletlike nanoparticles: mimicking shape, flexibility, and surface biology of platelets to target vascular injuries. Synthesis and characterization of thrombin conjugated g-Fe2O3 magnetic nanoparticles for hemostasis. Organ repair, hemostasis, and in vivo bonding of medical devices by aqueous solutions of nanoparticles. Nanoparticles: structure, properties, preparation and behaviour in environmental media. Antibiotic eluting clay mineral (Laponite(R)) for wound healing application: an in vitro study. Antimicrobial drugs encapsulated in fibrin nanoparticles for treating microbial infested wounds. Antibacterial effects of curcumin: an in vitro minimum inhibitory concentration study. Role of pro-oxidants and antioxidants in the anti-inflammatory and apoptotic effects of curcumin (diferuloylmethane). Curcumin-encapsulated nanoparticles as innovative antimicrobial and wound healing agent. Bactericidal actions of a silver ion solution on Escherichia coli, studied by energy-filtering transmission electron microscopy and proteomic analysis. Molecular and biological characterization of fibroblast growth factor, an angiogenic factor which also controls the proliferation and differentiation of mesoderm and neuroectoderm derived cells. Accelerated wound healing through the incorporation of basic fibroblast growth factor-impregnated gelatin microspheres into artificial dermis using a pressure-induced decubitus ulcer model in genetically diabetic mice. Hyaluronan based porous nano-particles enriched with growth factors for the treatment of ulcers: a placebo-controlled study. Genetic engineering of human stem cells for enhanced angiogenesis using biodegradable polymeric nanoparticles. Programming stem cells for therapeutic angiogenesis using biodegradable polymeric nanoparticles. Sonic hedgehog intradermal gene therapy using a biodegradable poly(beta-amino esters) nanoparticle to enhance wound healing. Small molecule end group of linear polymer determines cell-type gene delivery efficacy. Silver nanoparticles in therapeutics: development of an antimicrobial gel formulation for topical use. Silver nanoparticles mediate differential responses in keratinocytes and fibroblasts during skin wound healing. Nanocrystalline silver: a systematic review of randomized trials conducted on burned patients and an evidence-based assessment of potential advantages over older silver formulations. Composite nano-titanium oxide-chitosan artificial skin exhibits strong wound-healing effect-an approach with anti-inflammatory and bactericidal kinetics. Wound healing activity of Origanum vulgare engineered titanium dioxide nanoparticles in wistar albino rats. Tunable physiologic interactions of adhesion molecules for inflamed cell-selective drug delivery. Nanotechnology promotes the full-thickness diabetic wound healing effect of recombinant human epidermal growth factor in diabetic rats. Dual action antimicrobials: nitric oxide release from quaternary ammonium-functionalized silica nanoparticles. Shape- and nitric oxide flux-dependent bactericidal activity of nitric oxide-releasing silica nanorods. Nitric oxide-releasing quaternary ammonium-modified poly(amidoamine) dendrimers as dual action antibacterial agents. Nanoparticle (star polymer) delivery of nitric oxide effectively negates Pseudomonas aeruginosa biofilm formation. Role of size and shape on biofilm eradication for nitric oxide-releasing silica nanoparticles. Nitrosoglutathione generating nitric oxide nanoparticles as an improved strategy for combating Pseudomonas aeruginosa-infected wounds. Nitric oxide releasing nanoparticles are [82] [83] [84] [85] [86] [87] [88] [89] [90] [91] [92] therapeutic for Staphylococcus aureus abscesses in a murine model of infection. Systemic and immunotoxicity of silver nanoparticles in an intravenous 28 days repeated dose toxicity study in rats. Cellular toxicity of various inhalable metal nanoparticles on human alveolar epithelial cells. Neurotoxicity and gene-expressed profile in brain-injured mice caused by exposure to titanium dioxide nanoparticles. Uptake and toxicity of copper oxide nanoparticles in cultured primary brain astrocytes. Expectations of the benefits of nanotechnology are positive and have spread through all academic sectors of medicine, the wider scientific community, and industry in a general way [1]. Concurrent with this growth, however, there are increasing environmental and human health concerns [8e11]. In the medical sector, extensive research activities are in progress to develop nanoparticles, which can be used as efficient carriers for drug delivery through the skin barrier. In contrast, in cosmetic products, particles are mostly required to remain on the skin surface to fulfill their beneficial effect [1,12]. Nanocarriers can translocate intact into the skin without being degraded, or else they can be degraded at or near the skin surface, and the incorporated therapeutic molecule that is released can penetrate the skin layers. For optimum topical delivery of drugs into the skin, it is essential that the carrier releases the encapsulated drug, which can be further absorbed through the skin layers and through subcutaneous structures that are involved in the disease [11,13]. These processes are known to weaken the inside-out skin barrier function as measured by transepidermal water loss, which is believed to result from a disorganization of the intercellular lipid lamellae [25]. Skin delivery is mainly focused on topical delivery to treat local skin conditions or transdermal drug delivery, which involves the delivery of drugs through skin layers into the systemic circulation. The global term percutaneous/dermal absorption describes the passage of compounds across the skin. The process is divided into three steps: penetration, the entry of substance into a particular skin layer; permeation, the penetration through one layer into another; and, finally, resorption, the uptake into the vascular system [28]. Topical or transdermal delivery offers several advantages over the conventional oral and intravenous dosage forms, such as prevention of first-pass metabolism, minimization of pain, and possible controlled release of drugs [29]. Skin is a unique barrier composed of several highly organized and heterogeneous layers, and it includes a number of appendages such as hair follicles, and sweat and sebaceous glands. Dermis: A hydrophilic layer irrigated by the blood circulation and composed of a gel in which a densely network of fibers (collagen and elastin) provides mechanical strength to skin.
Cheap feldene online amex
Although there are other routes from the cochlear nuclei to the inferior colliculus arthritis banana diet buy generic feldene 20mg on-line, with additional intermediate relays, all ascending auditory pathways converge onto the inferior colliculus. Before moving on to the response properties of auditory neurons, we should make several points: 1. Projections and brain stem nuclei other than the ones described contribute to the auditory pathways. Each cochlear nucleus receives input from just the one ear on the ipsilateral side; all other auditory nuclei in the brain stem receive input from both ears. This explains the clinically important fact that the only way by which brain stem damage can produce deafness in one ear is if a cochlear nucleus (or auditory nerve) on one side is destroyed. Response Properties of Neurons in the Auditory Pathway To understand the transformations of auditory signals that occur in the brain stem, we must first consider the nature of the input from the neurons in the spiral ganglion of the cochlea. Because most spiral ganglion cells receive input from a single inner hair cell at a particular location on the basilar membrane, they fire action potentials only in response to sound within a limited frequency range. After all, hair cells are excited by deformations of the basilar membrane, and each portion of the membrane is maximally sensitive to a particular range of frequencies. Neural signals can travel from the spiral ganglion to auditory cortex via numerous pathways. Here, a primary pathway is shown schematically (at left) and through brain stem cross sections. The graph represents the firing rate in response to sounds at different frequencies. This type of frequency tuning is seen in many neurons in each of the relays from cochlea to cortex. For instance, some cells in the cochlear nuclei are especially sensitive to sounds varying in frequency over time (think of the sound of a trombone as it slides from a low note to a high note). An important development in the superior olive is that cells receive input from cochlear nuclei on both sides of the brain stem. As discussed later, such binaural neurons are probably important for sound localization. You can probably hear sounds you have been ignoring, and you can selectively pay attention to different sounds occurring at the same time. We are usually bathed in an amazing diversity of sounds-from chattering people to cars to electrical noises to sounds generated within our own bodies-and our brain must be able to analyze just the important sounds while ignoring the noise. We cannot yet account for the perception of each of these sounds by pointing to particular neurons in the brain. However, most sounds have certain features in common, including intensity, frequency, and the location from which they emanate. This neuron is frequency-tuned and has its greatest response at the characteristic frequency. As a stimulus gets more intense, the basilar membrane vibrates with greater amplitude, causing the membrane potential of the activated hair cells to be more depolarized or hyperpolarized. As a result, the nerve fibers with which the hair cells synapse fire action potentials at greater rates. In addition, more intense stimuli produce movements of the basilar membrane over a greater distance, which leads to the activation of more hair cells. In a single auditory nerve fiber, this increase in the number of activated hair cells causes a broadening of the frequency range to which the fiber responds. The loudness we perceive is correlated with the number of active neurons in the auditory nerve (and throughout the auditory pathway) and with their firing rates. Stimulus Frequency, Tonotopy, and Phase Locking From the hair cells in the cochlea through the various nuclei leading to auditory cortex, most neurons are sensitive to stimulus frequency. Frequency sensitivity is largely a consequence of the mechanics of the basilar membrane because different portions of the membrane are maximally deformed by sound of different frequencies. Moving from the base to the apex of the cochlea, a progressive decrease occurs in the frequency that produces a maximal deformation of the basilar membrane. From the base to the apex of the cochlea, the basilar membrane resonates with increasingly lower frequencies. In the cochlear nucleus, there are bands of cells with similar characteristic frequencies; characteristic frequencies increase progressively from anterior to posterior. Nearby neurons have similar characteristic frequencies, and a systematic relationship exists between position in the cochlear nucleus and characteristic frequency. In other words, there is a map of the basilar membrane within the cochlear nuclei. Because of the tonotopy present throughout the auditory system, the location of active neurons in auditory nuclei is one indication of the frequency of the sound. However, frequency must be coded in some way other than the site of maximal activation in tonotopic maps, for two reasons. One reason is that these maps do not contain neurons with very low characteristic frequencies, below about 200 Hz. As a result, the site of maximal activation might be the same for a 50 Hz tone as for a 200 Hz tone, so there must be some other way to distinguish them. At a fixed frequency, a more intense sound will produce a maximal deformation at a point further up the basilar membrane than a less intense sound. The timing of neural firing provides an additional type of information about sound frequency that complements information derived from tonotopic maps. If you think of a sound wave as a sinusoidal variation in air pressure, a phase-locked neuron would fire action potentials at the peaks, or the troughs, or some other constant location on the wave. Sound at a low frequency can elicit a phase-locked response, either (a) on every cycle of the stimulus or (b) on some fraction of the cycles. If you have a group of such neurons, each responding to different cycles of the input signal, it is possible to have a response to every cycle (by some member of the group) and thus a measure of sound frequency. It is likely that intermediate sound frequencies are represented by the pooled activity of a number of neurons, each of which fires in a phase-locked manner; this is called the volley principle. In other words, the sound waves cycle too fast for the action potentials of single neurons to accurately represent their timing. Many auditory neurons in the brainstem have peculiar membrane properties that make them uniquely sensitive to the precise timing of their synaptic inputs. Adaptations for precise timing are particularly impressive in neurons of the cochlear nuclei, as research by Donata Oertel and her colleagues at the University of Wisconsin has demonstrated (Box 11. To summarize, here is how different sound frequencies are represented by brain stem neurons. At very low frequencies, phase locking is used; at intermediate frequencies, both phase locking and tonotopy are useful; and at high frequencies, tonotopy must be relied on to indicate sound frequency. Neurons in the auditory nuclei of the brain stem can fire with a temporal precision better than 200 sec. Contrast this with cortical neurons, whose responses to identical stimuli are a hundred-fold less precise. In the auditory system, the timing of firing conveys important sensory information about the pitch of sounds and whether sounds come from the right or left. In the mid-1960s, computers made possible detailed analyses of the relationship between the waveforms of sounds and the firing of neurons. These studies, some done by my colleagues at the University of Wisconsin, revealed that auditory neurons encode the frequency of sounds not only by their position in the tonotopic map but also by firing in phase with the sounds, by phase locking. This temporal code breaks down at sound frequencies higher than 5 kHz because the firing of neurons is not precise enough to resolve periods shorter than about 200 sec. First, our impressive ability to distinguish tones as similar as 1000 Hz and 1002 Hz seems to depend on phase locking by neurons in the brain stem. Phase-locked neurons also detect the relative time of arrival of a sound at the two ears with every cycle of the sound, a mechanism important for localizing sounds in the horizontal plane. How can auditory neurons convey information with a temporal precision of 200 sec across multiple synapses- hair cells to spiral ganglion cells, to neurons of the cochlear nuclear nucleus, to neurons in the superior olive-using synaptic potentials and action potentials whose durations are in the millisecond range To achieve this, the firing of postsynaptic neurons must follow the firing of the presynaptic neurons rapidly and with an unvarying delay. In 1979, Bill Rhode, Phil Smith, and I began to address these issues by making intracellular recordings in anesthetized cats, but those experiments were terribly difficult.
Cheap feldene 20 mg without prescription
For example psoriatic arthritis in feet pictures buy 20mg feldene free shipping, the ability of testosterone to alter very young genitals and brain circuitry, leading to distinctly male genitals and masculine behaviors later in life, can be thought of as the organizational effects of the hormone. The hormone organizes the perinatal tissues in irreversible ways that allow it to generate male functions after sexual maturity has been reached. For a mature animal to express sexual behaviors fully, however, it is often necessary for steroid hormones to circulate again during periods of sexual activity, providing activational effects on the nervous system. Thus, for example, testosterone levels might surge in the body of a male songbird in the spring, activating changes in certain parts of his brain that are essential for normal reproductive behavior (Box 17. Steroids can directly affect transmitter synthesis, transmitter release, or postsynaptic transmitter receptors. The rise in testosterone that occurs prenatally actually leads to an increase in estrogen, which binds to estradiol receptors, thereby triggering masculinization of the developing nervous system. What is not clear is which genes are regulated in various parts of the brain by sex hormones to account for masculinization. Female gonads do not produce an early surge of testosterone or estrogen, so female brains are not affected in the same way as male brains. An interesting and important complication arises regarding the response of the fetal brain to circulating hormones. A reasonable question is why estrogens from the mother do not alter sexual development of the fetal brain. Singing is strictly a male function in many species, performed for the purpose of attracting and keeping a mate and for warning off potential rivals. Studies of two bird species with different habits of reproduction and singing have revealed some fascinating clues about the control and diversity of sexual dimorphisms in the brain. Zebra finches, which are popular pets, live in the wild in the harsh Australian desert. To breed successfully, birds require dependable sources of food, but in the desert, food comes only with sporadic and unpredictable rains. Zebra finches must therefore be ready and willing to breed whenever food and a mate are available, in any season. Wild canaries, on the other hand, live in the more predictable environment of the Azores and (where else They breed seasonally during spring and summer and do not reproduce during fall and winter. The males of both species are passionate singers, but they differ greatly in the size of their repertoires. Zebra finches belt out one simple ditty all their lives and cannot learn new ones. The different behaviors of zebra finches and canaries require different mechanisms of neural control. Birds sing by forcing air past a special muscularized organ called the syrinx, which encircles the air passage. However, the very different seasonal requirements of zebra finches and canaries are paralleled by distinctly different modes of steroidal control. Females that are not exposed to steroids when young are unresponsive to testosterone as adults. In contrast, the song system in canaries seems to be independent of early steroid exposure, yet it bursts into full service each spring. If female canaries are given androgens for the first time as adults, they will begin singing within a few weeks. By fall, male androgen levels drop, and the canary song system shrinks in size as his singing abates. In a sense, the male canary rebuilds much of his song control system anew each year as courtship begins. This may enable him to learn new songs more easily and, with his enlarged repertoire, gain some advantage in attracting a mate. As odd as it sounds, the female fetus must be protected from "female hormones" to keep its brain from becoming more masculine. Knockout mice that lack -fetoprotein are sterile and do not exhibit normal sexual behaviors. The role of -fetoprotein in humans is less clear; there are conflicting reports about whether human -fetoprotein binds estrogen as it does in rodents. Also noteworthy is the diagnostic measurement of -fetoprotein levels in maternal blood or amniotic fluid; unusually high levels are a possible indicator of neural tube defects, and unusually low levels are found in Down syndrome. A study by Amateau and McCarthy shows that one factor in brain masculinization "downstream" from testosterone and estradiol is the production of prostaglandins. Prostaglandins are compounds derived from arachidonic acid, a fatty acid present in the brain and other organs. Prostaglandins have numerous roles; most notably, they are produced after tissue damage and are involved in inducing pain and fever. Under normal circumstances, the genetic sex of an animal or person determines hormonal function and, consequently, the sexual characteristics of the nervous system. However, in situations in which hormonal function is altered, it is possible for genetic males to have female brains and genetic females to have male brains. For example, in all mammalian species studied, treatment with testosterone early in development leads to decreases in at least some features of adult female sexual behavior. Activating fully masculine behavior usually requires extended testosterone treatment before and after birth. Female guinea pigs treated in utero with enough testosterone to masculinize their external genitals will, as adults, energetically mount and attempt to mate with females in estrus. In the absence of human intervention, when a cow carries twin calves that include both a male and a female, the female calf is exposed in utero to some testosterone produced by her male twin. As an adult, the female, known as a freemartin, will invariably be infertile and behave more like a bull than a cow. Some humans also experience mismatches between their chromosomes and sex hormones. The androgen receptor gene is on the X chromosome; males thus have only one copy of it, and males with the defective gene cannot produce functioning androgen receptors. These individuals develop normal testes but they remain undescended in the abdomen. Androgen-insensitive genetic males not only look like normal genetic females, but they also behave like them. Even when they understand the circumstances of their biology, they prefer to call themselves women; they dress like women, and they choose men as their sex partners. At birth, they have normal ovaries and no testes, but their external genitals are intermediate in size between a normal clitoris and a penis. We have to be particularly cautious about drawing conclusions about the causes of human behavior, however (Box 17. While there is no question that hormones are extremely important in sexual development, recent research suggests that genes may sometimes be more directly involved in sexual differentiation, at least in some species. As both sides of the brain are exposed to the same circulating hormones, they should be equally masculine or feminine if hormones are entirely responsible for sexual differentiation. In related studies of gynandromorphic chickens, researchers found that most cells on the male side of the body carry male sex chromosomes and cells on the other side have female chromosomes, the result of a mutation early in development. Recently, attempts have been made to find direct genetic effects on sexual differentiation in mammals. But circumcision, an accident with an electrocautery device burned his entire penis. It also casts doubt on the theory that major sex differences, psychological as well as anatomical, are immutably set by the genes at conception. Despite cosmetic surgery and female indoctrination, as an adult, he said that he had suspected he was a boy as early as the second grade and imagined growing up to be a muscular man. He knew nothing about the failed circumcision and subsequent surgery, nor the fact that he was genetically a male. For years, David dealt with the overwhelming emotional problems resulting from his past. Tragically, after numerous traumatic events in his life, including the death of his twin brother and the breakup of his marriage, David committed suicide in 2004 at the age of 38.
Tibetan Ginseng (Ginseng, Panax). Feldene.
- What is Ginseng, Panax?
- Premature ejaculation when a cream containing ginseng and other ingredients is applied directly to the skin of the penis.
- Dosing considerations for Ginseng, Panax.
- Are there any interactions with medications?
- Male impotence (erectile dysfunction).
Source: http://www.rxlist.com/script/main/art.asp?articlekey=96961
Order generic feldene on line
Consequently arthritis physical therapy order 20mg feldene with visa, the male montane voles had numbers of vasopressin receptors in the ventral pallidum comparable to prairie voles. If this cause-and-effect link is supported by further studies, it will dramatically show that a complex social behavior can be altered by the overexpression of a single protein at one location in the brain. These coronal brain sections show the distribution of oxytocin and vasopressin in the brains of montane voles and prairie voles. The research on voles suggests a very interesting hypothesis about the evolution of complex social behaviors. Love, Bonding, and the Human Brain the vole story is a fascinating example of how brain chemicals can regulate critical behaviors. But what does all this have to do with human relationships, faithfulness, and love It is too early to be sure, but intriguing pieces of evidence suggest that voles may teach us something about the human brain and behavior. For example, there is evidence that human plasma oxytocin levels increase during breastfeeding in mothers and during sexual intercourse in men and women. In an experiment investigating maternal love, while brain scans were being taken, mothers saw pictures of their child intermixed with pictures of other familiar children. In a second experiment studying romantic love, brain activity was compared when men and women viewed pictures of partners and pictures of friends. Brain activation is shown in (a) sagittal, (b) horizontal, and (c,d) two different coronal planes. Yellow areas were more active when mothers saw pictures of their own child than with pictures of other familiar children. Red areas were more activated by pictures of romantic partners than to pictures of friends. We can speculate that the brain activation demonstrates the strong reinforcing nature of partner and parental relationships. Although human behavior undoubtedly involves more complex factors than vole behavior, surprising evidence suggests that vasopressin influences human bonding as well. Hasse Walum and a team of Swedish and American scientists studied 552 pairs of same-sex Swedish twins who were married or had long-term partners. Of particular interest was the gene sequence that codes for vasopressin receptors and the tendency toward monogamy. When this gene variant is transgenically introduced into nonmonogamous mice, their social behavior becomes more like the prairie voles. In the human twin study, the scientists investigated whether vasopressin gene variants might influence pairbonding in humans too. In women, there was no connection between the vasopressin gene variants and the quality of their marriage as assessed by a variety of questionnaires. In the men, however, an intriguing correlation was found: Men with a particular gene variant scored significantly lower on measures of the quality of their marriage and were twice as likely to report that a marital crisis had occurred in the year before the survey. The wives of the men with the gene variant also reported lower marital quality than the wives of the men without the variant. The function of this gene variant is not known, but these results suggest that, even in humans, vasopressin receptors might play a role in pair-bonding. Since all behaviors ultimately depend on the structure and function of the nervous system, we can make the strong prediction that male and female brains are also somehow different; that is, they have sexual dimorphisms (from the Greek dimorphos, "having two forms"). Another good reason to expect that male and female brains differ is simply that male and female bodies differ. The body parts that are unique to each sex require neural systems that have evolved specifically to control them. For example, male rats have a particular muscle at the base of the penis, and their spinal cord has a small cluster of motor neurons that control that muscle; females lack both the muscle and the related motor neurons. Body size and general shape also vary with gender, and thus somatosensory and motor maps must adjust to fit them. These brains are from adult female (left) and male (right) three-spined stickleback fish that were the same length and weight. In rodents, the trained eye can tell male from female brains with no ambiguity because of differences in the hypothalamus. The diversity of brain dimorphisms across species is sometimes associated with remarkable variations in sexual behaviors. For example, in some songbird species, only males sing, and, not surprisingly, only males have large singing-related brain nuclei. In human brains, dimorphisms have so far proven to be subtle, few, and of unknown function. Differences between human male and female brains tend to vary along a continuum, with lots of overlap. A particular hypothalamic nucleus might be larger in women than men on average, for example, but size variations of the nucleus may be so great that many men have a larger nucleus than many women. In the rest of this section, we will describe sexual dimorphisms in the nervous systems of humans and other species, focusing on examples that illuminate the relationship between the brain and behavior. We will also discuss some of the neurobiological mechanisms that generate these dimorphisms. Sexual Dimorphisms of the Central Nervous System Few dimorphic neural structures are related to their sexual functions in an obvious way. In women, it surrounds the opening of the vagina and serves to constrict it slightly. The most distinct sexual dimorphisms in the mammalian brain are clustered around the third ventricle, within the preoptic area of the anterior hypothalamus. The preoptic area of humans may also have dimorphisms, but the differences are small and controversial. Various neurons of the medial preoptic area in male rhesus monkeys fire vigorously during specific phases of sexual behavior, including arousal and copulation. In addition, there may be subtle differences in the size of certain hypothalamic nuclei that correlate with sexual orientation in people. Human brain dimorphisms outside the hypothalamus have been difficult to prove conclusively, although many have been reported. For example, some studies have found that the corpus callosum is larger in men, but this may be a consequence of men having slightly larger brains (and bodies). In other reports, the posterior end of the corpus callosum, called the splenium, is larger in women than men. But even if there is a dimorphism in the size or shape of the corpus callosum, what could it mean The callosum has no obvious role in mediating specifically sexrelated behaviors, but it is important for a variety of cognitive functions that involve coordinated activity between hemispheres. Observations of stroke patients in whom only one hemisphere has been damaged suggest that the functions of female brains may be less lateralized, that is, less dependent on one cerebral hemisphere more than the other. Perhaps, the most reliable conclusion we can draw about sexual dimorphisms in human brain structure is that there are so few of them. The gross anatomy of the brain provides only a crude view of the organization of the nervous system. To determine the reasons for sexually dimorphic behavior, we need to look deeper into the patterns of neural connections, the neurochemistry of the brain, and the influence of sex-related hormones on neural development and function. Sexual Dimorphisms of Cognition Even if there are no major differences in the brain structures of men and women, there may still be differences in cognitive abilities. Reports of cognitive dimorphisms are sometimes accompanied by an evolutionary explanation: Men evolved as hunters and relied more on their abilities to navigate their environment. Women evolved the behavior of staying closer to home to care for children, so they were more social and verbal. Starting at around age 11 years, girls perform slightly better on tests of comprehension and writing, and this effect is sometimes said to extend through high school and beyond. Perhaps, it reflects a difference in the rates of brain development in the two sexes. Tasks that reportedly favor men include map reading, maze learning, and mathematical reasoning. Researchers speculate that these male advantages evolved from the days when men roamed large areas to hunt wild animals. Second, across large groups of people of both sexes, there are huge differences in performance.
Discount feldene online mastercard
If a child is born cross-eyed and the condition is not corrected before the age of 10 years zen arthritis cream buy generic feldene 20 mg online, binocular depth perception will be lost forever. From your knowledge of the central visual system, where do you think the circuitry has been modified What sort of experiment might you perform to investigate the relationship between visual perception and neural activity in the visual cortex Hearing is a vivid part of our conscious lives, while balance is something we experience all day but rarely think about. When we cannot see something or someone, we can often detect its presence, identify its origin, and even receive a message from it just by hearing its sounds. Aside from the ability to detect and locate sound, we can perceive and interpret its nuances. We can immediately distinguish the bark of a dog, the voice of a particular friend, the crash of an ocean wave. Because humans are able to produce a wide variety of sounds as well as hear them, spoken language and its reception via the auditory system have become an extremely important means of communication. Audition in humans has even evolved beyond the strictly utilitarian functions of communication and survival; musicians, for example, explore the sensations and emotions evoked by sound. In contrast to hearing, the sense of balance is strictly a personal, internalized process. The vestibular system informs our nervous system where our head and body are and how they are moving. This information is used, without conscious effort, to control muscular contractions that keep or put our body where we want it to be, to reorient ourselves when something moves us, and to move our eyes so that our visual world stays fixed on our retinas even when our head is bouncing around. Here, we will explore the mechanisms within the ear and brain that translate the sounds in our environment into meaningful neural signals and the movements of our head into a sense of where we are. We will find that these transformations are carried out in stages rather than all at once. Within the inner ear, neural responses are generated by auditory receptors from the mechanical energy in sound and by vestibular receptors from the tilts and rotations of the head. At subsequent stages in the brain stem and thalamus, signals from the receptors are integrated before they ultimately reach auditory and vestibular cortex. By looking at the response properties of neurons at various points in the system, we begin to understand the relationship between neural activity and our perception of sound and balance. Almost anything that can move air molecules can generate a sound, including the vocal cords of the human larynx, the vibration of a string on a guitar, and the explosion of a firecracker. When an object moves toward a patch of air, it compresses the air, increasing the density of the molecules. These changes in air pressure are transferred away from the speaker at the speed of sound, which is about 343 m/sec (767 mph) for air at room temperature. When the paper cone of a stereo speaker pushes out, it compresses the air; when the cone pulls in, it rarefies the air. If the push and pull are rhythmic, there will also be a rhythmic variation in the air pressure, as shown in the graph. The distance between successive compressed (high-pressure) patches of air is one cycle of the sound (indicated by the vertical lines). The frequency of the sound is the number of compressed or rarefied patches of air that pass by our ears each second. One cycle of the sound is the distance between successive compressed patches; the sound frequency, expressed in units called hertz (Hz), is the number of cycles per second. Each graph plots air pressure versus distance for a sound of constant frequency and intensity. Note that the X axis also represents time because the velocity of sound is constant. Whether a sound is perceived to have a high or low tone, or pitch, is determined by the frequency. In order to understand frequency, realize that a room-shaking low note on an organ is about 20 Hz and an ear-piercing high note on a piccolo is about 10,000 Hz. Although humans can hear a great range of frequencies, there are high and low sound wave frequencies our ears cannot hear, just as there are electromagnetic waves of light our eyes cannot see (Box 11. Sound intensity determines the loudness we perceive, loud sounds having higher intensity. If our auditory system were much more sensitive, we would hear a constant roar from the random movement of air molecules. Real-world sounds rarely consist of simple rhythmic sound waves at one frequency and intensity. It is the simultaneous combination of different frequency waves at different intensities that gives different musical instruments and human voices their unique tonal qualities. For instance, ultrasonic dog whistles work because dogs can hear up to about 45 kHz. Some bats vocalize at frequencies up to 100 kHz, then listen to the echoes of their calls in order to locate objects (see Box 11. Some fish in the shad and herring family can detect sounds as high as 180 kHz, thereby enabling them to hear the echolocating ultrasound generated by dolphins that prey on them. Similarly, nocturnal moths listen for the ultrasound of hungry bats, so they can evade these predators. Some animals can hear infrasonic frequencies; one is the elephant, which can detect 15 Hz tones inaudible to humans. Whales produce low-frequency sounds, which are thought to be a means of communication over distances of many kilometers. The earth also produces low-frequency vibrations, and some animals may sense an impending earthquake by hearing such sounds. Even though we usually cannot hear very low frequencies with our ears, they are present in the environment and we can sometimes feel them as vibrations with our somatosensory system (see Chapter 12). Infrasound is produced by such devices as air conditioners, boilers, aircraft, and automobiles, and it can have unpleasant subconscious effects. Although even intense infrasound from these machines does not cause hearing loss, it can cause dizziness, nausea, and headache. At very high levels, low-frequency sound may also produce resonances in body cavities such as the chest and stomach, which can damage internal organs. In addition to mechanical equipment, our own bodies generate inaudible low-frequency sound. When muscle changes length, individual fibers vibrate, producing low-intensity sound at about 25 Hz. While we cannot normally hear these sounds, you can demonstrate them to yourself by carefully putting your thumbs in your ears and making a fist with each hand. As you tighten your fist, you can hear a low rumbling sound produced by the contraction of your forearm muscles. Other muscles, including your heart, produce inaudible sound at frequencies near 20 Hz. The visible portion of the ear consists primarily of cartilage covered by skin, forming a sort of funnel called the pinna (from the Latin for "wing"), which helps collect sounds from a wide area. The shape of the pinna makes us more sensitive to sounds coming from ahead than from behind. In humans, the pinna is more or less fixed in position, but animals such as cats and horses have considerable muscular control over the position of their pinna and can orient it toward a source of sound. The entrance to the internal ear is called the auditory canal, which extends about 2. Connected to the medial surface of the tympanic membrane is a series of bones called ossicles (from the Latin for "little bones"; the ossicles are indeed the smallest bones in the body). Located in a small air-filled chamber, the ossicles transfer movements of the tympanic membrane into movements of a second membrane covering a hole in the bone of the skull called the oval window. Behind the oval window is the fluid-filled cochlea, which contains the apparatus for transforming the physical motion of the oval window membrane into a neuronal response. Thus, the first stages of the basic auditory pathway look like this: Sound wave moves the tympanic membrane. The structures from the pinna to the tympanic membrane make up the outer ear, the tympanic membrane and the ossicles constitute the middle ear, and the apparatus medial to the oval window is the inner ear.
Proven 20mg feldene
Our progress rested on the diverse expertise and hard work of these collaborators arthritis of the spine 20 mg feldene fast delivery. I believe that a team approach is an absolute necessity for tackling the extraordinarily complex functions and malfunctions of the central nervous system. Working collaboratively exposes us to new ideas and promotes our growth as scientists. Permanent central synaptic disconnection of proprioceptors after nerve injury and regeneration. Imagine a situation in which muscle contraction is commanded by an upper motor neuron. The alpha motor neurons respond, the extrafusal fibers contract, and the muscle shortens. If they were to become slack, the Ia axons would become silent and the spindle would go "off the air," no longer providing information about muscle length. This does not happen, however, because the gamma motor neurons are also activated. Gamma motor neurons innervate the intrafusal muscle fiber at the two ends of the muscle spindle. Activation of these fibers causes a contraction of the two poles of the muscle spindle, thereby pulling on the noncontractile equatorial region and keeping the Ia axons active. Notice that the activation of alpha and gamma motor neurons has opposite effects on Ia output; alpha activation alone decreases Ia activity, while gamma activation alone increases Ia activity. Recall from our discussion earlier that the monosynaptic stretch reflex arc can be viewed as a feedback loop. The principles of feedback control systems are that a set point is determined (in this case, the desired muscle length), deviations from the set point are detected by a sensor (the Ia axon endings), and deviations are compensated for by an effector system (alpha motor neurons and extrafusal muscle fibers), returning the system to the set point. This circuit, gamma motor neuron intrafusal muscle fiber Ia afferent axon alpha motor neuron extrafusal muscle fibers, is sometimes called the gamma loop. During most normal movements, alpha and gamma motor neurons are simultaneously activated by descending commands from the brain. By regulating the set point of the stretch feedback loop, the gamma loop provides additional control of alpha motor neurons and muscle contraction. Muscle Proprioception from Golgi Tendon Organs Muscle spindles are not the only source of proprioceptive inputs from the muscles. Another sensor of skeletal muscle is the Golgi tendon organ, which acts like a very sensitive strain gauge; that is, it monitors muscle tension, or the force of contraction. They are located at the junction of the muscle and the tendon and are innervated by group Ib sensory axons, which are slightly smaller than the Ia axons innervating the muscle spindles. As the fibrils straighten and squeeze the Ib axons, their mechanosensitive ion channels are activated and action potentials can be triggered. This different anatomical arrangement helps to determine the types of information these two sensors provide the spinal cord: Ia activity from the spindle encodes muscle length information, while Ib activity from the Golgi tendon organ encodes muscle tension information. The Ib axons enter the spinal cord, branch repeatedly, and synapse on special interneurons called Ib inhibitory interneurons in the ventral horn. Ib interneurons also receive inputs from other sensory receptors and from descending pathways. Because the activated muscle does not change length, the Ia axons remain silent in this example. The Ib axon of the Golgi tendon organ excites an inhibitory interneuron, which inhibits the alpha motor neurons of the same muscle. However, its normal function is to regulate muscle tension within an optimal range. As muscle tension increases, the inhibition of the alpha motor neuron slows muscle contraction; as muscle tension falls, the inhibition of the alpha motor neuron is reduced, and muscle contraction increases. This type of proprioceptive feedback is thought to be particularly important for the proper execution of fine motor acts, such as the manipulation of fragile objects with the hands, which require a steady, but not too powerful, grip. We have focused on the proprioceptors that are involved in reflex control of the spinal motor neurons. However, besides muscle spindles and Golgi tendon organs, a variety of proprioceptive axons are present in the connective tissues of joints, especially within the fibrous tissue surrounding the joints (joint capsules) and ligaments. These mechanosensitive axons respond to changes in the angle, direction, and velocity of movement in a joint. Most are rapidly adapting, meaning that sensory information about a moving joint is plentiful, but nerves encoding the resting position of a joint are few. We are, nevertheless, quite good at judging the position of a joint, even with our eyes closed. Removing one source of information can be compensated for by the use of the other sources. When an arthritic hip is replaced with a steel and plastic one, patients can still tell the angle between their thigh and their pelvis, despite the fact that all their hip joint mechanoreceptors are sitting in a jar of formaldehyde. Spinal Interneurons the actions of Ib inputs from Golgi tendon organs on alpha motor neurons are entirely polysynaptic; they are all mediated by intervening spinal interneurons. Indeed, most of the input to the alpha motor neurons comes from interneurons of the spinal cord. The interneurons are themselves networked together in a way that allows coordinated motor programs to be generated in response to their many inputs. Interneurons play a critical role in the proper execution of even the simplest reflexes. Compensation for the lengthening of one set of muscles, such as the flexors of the elbow, involves contraction of the flexors via the stretch reflex but also requires relaxation of the antagonist muscles, the extensors. This process is called reciprocal inhibition, the contraction of one set of muscles accompanied by the relaxation of their antagonist muscles. The importance of this is obvious; imagine how hard it would be to lift something by contracting your biceps if its antagonist muscles. Reciprocal inhibition is also used by descending pathways from the brain to overcome the powerful stretch reflex. Consider a situation in which the flexors of the elbow are voluntarily commanded to contract. You might expect the resulting stretch of the antagonist extensor muscles to activate their stretch reflex arc, which would strongly resist flexion of the joint. However, the descending pathways that activate the alpha motor neurons controlling the flexors also activate interneurons, which inhibit the alpha motor neurons that supply the antagonist muscles. This is a complex, polysynaptic reflex arc used to withdraw a limb from an aversive stimulus (such as the withdrawal of your foot from the thumbtack in Chapter 3). The direction of withdrawal depends on the location of the stimulus; for example, hot stimuli applied to your palm and to the back of your hand trigger withdrawals in opposite directions (as you would hope! The flexor reflex is far slower than the stretch reflex, indicating that a number of interneurons intervene between the sensory stimulus and the coordinated motor act. The flexor reflex is activated by the small, myelinated A nociceptive axons that trigger pain (see Chapter 12). The nociceptive axons entering the spinal cord branch profusely and activate interneurons in several different spinal segments. These cells eventually excite the alpha motor neurons that control all the flexor muscles of the affected limb (and, needless to say, inhibitory interneurons are also recruited to inhibit the alpha motor neurons that control the extensors). Luckily, an additional component of the reflex is recruited: the activation of extensor muscles and the inhibition of flexors on the opposite side. Notice that this is another example of reciprocal inhibition, but in this case, activation of the flexors on one side of the spinal cord is accompanied by inhibition of the flexors on the opposite side. The Generation of Spinal Motor Programs for Walking the crossed-extensor reflex, in which one limb extends as the other limb flexes, seems to provide a building block for locomotion. In principle, this could be a series of descending commands from upper motor neurons. However, as we already suspected from our consideration of headless chicken behavior, it seems likely that this control is exerted from within the spinal cord. The circuit for the coordinated control of walking must reside, therefore, within the spinal cord. In general, circuits that give rise to rhythmic motor activity are called central pattern generators. However, the simplest pattern generators are individual neurons whose membrane properties endow them with pacemaker properties. An interesting example comes from the work of Sten Grillner and his colleagues in Stockholm, Sweden. Based on the assumption that the spinal central pattern generators for locomotion in different species are variations on a plan that was established in a common ancestor, Grillner focused on the mechanism for swimming in the lamprey, a jawless fish that has evolved slowly over the course of the past 450 million years.
Buy feldene master card
Most studies rely on the use of invasive sampling methods such as biopsied skin or excised human skin [7 arthritis in dogs hips treatment buy generic feldene 20 mg,34e36] or animal models [37e39]. The extensive use of these ex vivo methods in industry and academic research is tied to benefits like having fewer regulatory issues to overcome and less time-consuming experiments. The accuracy of these in vitro experiments, on the other hand, relies strongly on study approach in terms of skin model, experimental setup, and analytical method [33]. In vivo assessment on human volunteers for defining nanoparticle penetration remains the most relevant model but is challenging due to health, safety, and ethical reasons. Noninvasive imaging techniques provide real-time risk assessment on nanoparticle exposure and penetration in humans without requiring invasive sampling. Here, we discuss some of the imaging technologies that are commonly used to localize nanoparticles in human volunteer skin. This has attracted researchers to apply it to a wide range of biological applications, including assessment of skin conditions [44,45], in vivo cell-trafficking tracking [46], metabolic imaging [47], drug delivery [22], and stem cell optoinjection [48]. Yet the application of this technology has been limited by the substantial acquisition (w300,000V) and maintenance cost of the instrument due to the need for a femtosecond excitation laser source. The fact that nanotechnologies are a fast-growing market and are already in use in a variety of consumer products means that people are increasingly exposed to newly developed nanomaterials. A number of studies have shown that nanoparticles can be toxic to living cells [52,53], and the question of whether existing approaches can accurately assess risks of these nanoparticles to human health remains debatable. One of the fundamental strategies to assess this risk is by determining whether the nanoparticles breach the skin barrier and reach viable tissue. This technique also has the ability to characterize the elemental component of the nanoparticles. The excitation beam (red) passes through a dichroic mirror and optical lenses before reaching the sample. The emitted light (blue) then passes back to the lenses and is reflected to one or more detectors by the dichroic mirror. In the case of examining a complex medium, for instance nanoparticles within skin, a doubleexponential decay profile can be applied during data processing. These numerical outputs can be analyzed to differentiate nanoparticle signals from endogenous signals. However, the nanoparticles need to be intrinsically fluorescent or have fluorescent tags to be detected by this form of microscopy. Another problem with this technique is the risk of damage to both viable tissue and the fluorophore resulting in photobleaching [69]. However, many of these imaging technologies are seen as complementary and often are paired to improve assessment [70]. The typical configuration of a single-photon confocal microscope includes a laser light source to illuminate a fluorescent derivative, a condenser, objective lenses, and a point detector. Structures in the grayscale reflectance images that appear bright have components with a high reflective index compared to the surrounding tissue. Silver nanoparticle aggregates (white arrowheads) were observed within the skin furrows at 2 and 6 days (data not shown) after application in intact and tape-stripped skin. No aggregates were seen after 10 days in intact skin, but discrete aggregates were observed in tape-stripped skin. We detected no aggregates in intact skin 10 days post treatment, while the aggregates in tapestripped skin appeared to persist on the surface of skin, within furrows, and in hair follicles. In brief, the technology uses an interferometry technique to collect structural data from complex media such as skin. A reference profile is generated from the reflectivity from the mirror path and is compared to the sample path. Lateral scanning (open double arrow) is performed by the moving focus of the probing beam and directed to the sample after focusing through the galvanometer scanning mirrors and optics. Depth data (A-scan) are obtained by depth shifting the coherence window with the reference mirror (A). Scattered pigment can be located at the subepidermal level based on the en-face image. The white arrows in (B) indicate bright structures with a dark center that may correspond to pigment-loaded macrophages. Similar to most noninvasive imaging technologies, imaging can be done without the need for excision or tissue processing. The presence of the nodular granuloma containing lymphohistiocytic infiltrates was further confirmed by histopathological findings from a skin biopsy. One of the first questions to determine whether a particular nanoparticle causes toxicity is to find out if the particle can overcome the skin barrier and penetrate living tissues of the skin. Current methodologies to investigate skin penetration mostly consist of invasive techniques that make human studies difficult and can involve substantial amounts of tissue processing. Noninvasive imaging provides an alternative and complementary method for more accurate assessments of nanotoxicity in the whole, as the study can be carried out in human volunteers with minimal issues. This form of analysis also allows for longitudinal studies of nanotoxicity, which are impossible with destructive analytical approaches. That said, there are several limitations to consider before imaging nanoparticles in skin. One limitation of imaging nanoparticles in skin is the vast difference in the size of skin features and the diameter of nanoparticles; for example, skin features are measured in micrometers, whereas nanoparticle features are measured in nanometers. The size of a single nanoparticle is at least 30,000 times smaller than a single keratinocyte [78]; one practical limitation is that when we image different skin strata, with hematoxylin and eosin (H&E)-stained sections for instance, we will not be able to visualize the nanoparticles with the same light microscope due to insufficient magnification and resolution. One is to use electron microscopy to study individual nanoparticles at subcellular resolution, with the limitations of analyzing only a small area and the risk of overinterpreting artifacts. Another is the use of noninvasive imaging techniques to assess the skin organization, disregarding probable errors in the number of nanoparticles and their location. The ideal approach for studying toxicity of a nanoparticle would be to combine high-resolution imaging in excised human skin and noninvasive imaging technologies in human volunteers. The ability to differentiate what is and is not a nanoparticle is one of the most challenging aspects of studying nanoparticles in skin. For example, the actual concentration exposed to skin in reality may be below the detectable limit of the imaging tool available, but that does not conclude that there are no nanoparticles that had penetrated the skin. It is also possible that a researcher might not visualize the nanoparticles the same way others reported in their studies. We routinely image high concentrations of nanoparticles blotted onto paper as a positive control when setting up a new experiment. This approach helps us to more accurately interpret the imaging data and avoid artifacts. The use of these imaging instruments becomes increasingly intricate as the complexity of these machines increases. Each imaging modality has pros and cons in terms of imaging nanoparticles in skin and potential artifacts. Multimodality imaging combines the strengths of individual modalities while 364 27. We believe that integrating multiple imaging modalities can improve skin penetration data and provide more accurate and comprehensive information for diagnosis of diseases. A predictive algorithm for skin permeability: the effects of molecular size and hydrogen bond activity. Estimation of physicochemical parameters, and application to a large permeability database. Relative uptake of minoxidil into appendages and stratum [21] [22] [23] [24] [25] [26] [27] [28] [29] [30] [31] corneum and permeation through human skin in vitro. Hair follicles contribute significantly to penetration through human skin only at times soon after application as a solvent deposited solid in man. A review of the scientific literature on the safety of nanoparticulate titanium dioxide or zinc oxide in sunscreens. The human stratum corneum layer: an effective barrier against dermal uptake of different forms of topically applied micronised titanium dioxide. Determination of the cuticula thickness of human and porcine hairs and their potential influence on the penetration of nanoparticles into the hair follicles. Potential use of nanoparticles for transcutaneous vaccine delivery: effect of particle size and charge.
