

Order 250mg diamox with visa
In particular symptoms qt prolongation generic diamox 250 mg with visa, the theoretical predictions about engulfment patterns of vesicles with nonspherical shapes, as described in Section 8. Vesicle membrane segments corresponding to the free, Fst, or bistable, B st, regimes should appear dark, whereas completely engulfed, Cst, segments should show a homogeneous fluorescence. For membrane segments that belong to a partially engulfed, Pst, regime, curvature-mediated interactions will lead to more complex behavior. Such experimental studies of engulfment patterns would also provide a direct test of the stability relations in Eqs. Furthermore, using partially adhesive Janus-like particles in contact with deflated vesicles of nonspherical shape, it should be possible to experimentally study the curvature-induced forces described in Section 8. It will also be interesting to directly probe the bistable regime, Bst, in which both the free and completely engulfed particle states are (meta)stable, using experiments with optical tweezers. Indeed, we expect that, in a bistable regime, a particle in contact with the membrane will not be spontaneously engulfed. However, if the particle is pushed against the membrane by the tweezers, it should become completely engulfed, and it should remain engulfed even if the force exerted by the tweezers is switched off. In order to be released, the particle will have to be pulled out of the vesicle using the tweezers once again. Other experimental challenges that will prove interesting, and which will require an expansion of the theoretical ideas described in this chapter, involve the interaction of particles with more complex vesicles, such as (i) vesicles with phase-separated membranes and (ii) vesicles with nanotubes, both described in Chapter 5. In the case of phase-separated vesicles, the particles should be preferentially engulfed by liquid-disordered domains rather than by liquid-ordered domains, because liquid-disordered membranes have a lower bending rigidity, unless this effect is overcompensated by a stronger adhesion of the particles to the liquid-ordered domains. Furthermore, the positive line tension of the domain boundaries should favor particle engulfment, because the engulfment process will tend to reduce the length of the contact line between domains. In the case of vesicles with nanotubes, the nanotubes should act as an area reservoir that can be used to engulf particles if these are sufficiently adhesive in the same way that neutrophils can extract area from membrane reservoirs during phagocytosis of many large particles (Herant et al. Whereas spherical vesicles without nanotubes cannot engulf particles without having their membrane stretched and ruptured, see Section 8. Agudo-Canalejo J, Lipowsky R (2015b) Adhesive nanoparticles as local probes of membrane curvature. Agudo-Canalejo J, Lipowsky R (2016) Stabilization of membrane necks by adhesive particles, substrate surfaces, and constriction forces. Agudo-Canalejo J, Lipowsky R (2017) Uniform and Janus-like nanoparticles in contact with vesicles: Energy landscapes and curvature-induced forces. Dasgupta S, Auth T, Gompper G (2013) Wrapping of ellipsoidal nanoparticles by fluid membranes. Dasgupta S, Auth T, Gompper G (2014) Shape and orientation matter for the cellular uptake of nonspherical particles. Gruhn T, Franke T, Dimova R, Lipowsky R (2007) Novel method for measuring the adhesion energy of vesicles. Guo R, Mao J, Yan L-T (2013) Unique dynamical approach of fully wrapping Dendrimer-like soft nanoparticles by lipid bilayer membrane. Herant M, Heinrich V, Dembo M (2005) Mechanics of neutrophil phagocytosis: Behavior of the cortical tension. Huang C, Zhang Y, Yuan H, Gao H, Zhang S (2013) Role of nanoparticle geometry in endocytosis: Laying down to stand up. Jaskiewicz K, Larsen A, Schaeffel D, Koynov K, Lieberwirth I, Fytas G, Landfester K, Kroeger A (2012) Incorporation of nanoparticles into polymersomes: Size and concentration effects. Kusumaatmaja H, Lipowsky R (2011) Droplet-induced budding transitions of membranes. Li S, Malmstadt N (2013) Deformation and poration of lipid bilayer membranes by cationic nanoparticles. Li Y, Li X, Li Z, Gao H (2012) Surface-structure-regulated penetration of nanoparticles across a cell membrane. Liu J, Lu N, Li J, Weng Y, Yuan B, Yang K, Ma Y (2013) Influence of surface chemistry on particle internalization into giant unilamellar vesicles. Luccardini C, Tribet C, Vial F, Marchi-Artzner V, Dahan M (2006) Size, charge, and interactions with giant lipid vesicles of quantum dots coated with an amphiphilic macromolecule. Mai Y, Eisenberg A (2010) Controlled incorporation of particles into the central portion of vesicle walls. Michel R, Kesselman E, Plostica T, Danino D, Gradzielski M (2014) Internalization of silica nanoparticles into fluid liposomes: Formation of interesting hybrid colloids. Noguchi H, Takasu M (2002) Adhesion of nanoparticles to vesicles: A Brownian dynamics simulation. Pan Y, Neuss S, Leifert A, Fischler M, Wen F, Simon U, Schmid G, Brandau W, Jahnen-Dechent W (2007) Size-dependent cytotoxicity of gold nanoparticles. Panyam J, Labhasetwar V (2003) Biodegradable nanoparticles for drug and gene delivery to cells and tissue. Rai M, Yadav A, Gade A (2009) Silver nanoparticles as a new generation of antimicrobials. Sari A, Cacciuto A (2012) Mechanism of membrane tube formation induced by adhesive nanocomponents. Schmid K, Riediker M (2008) Use of nanoparticles in Swiss industry: A targeted survey. Seifert U, Berndl K, Lipowsky R (1991) Shape transformations of vesicles: Phase diagram for spontaneous-curvature and bilayercoupling models. Verma A, Stellacci F (2010) Effect of surface properties on nanoparticlecell interactions. Wei X, Jiang W, Yu J, Ding L, Hu J, Jiang G (2015) Effects of SiO2 nanoparticles on phospholipid membrane integrity and fluidity. Yang K, Ma Y-Q (2010) Computer simulation of the translocation of nanoparticles with different shapes across a lipid bilayer. Yi X, Shi X, Gao H (2014) A universal law for cell uptake of onedimensional nanomaterials. Yue T, Zhang X, Huang F (2014) Membrane monolayer protrusion mediates a new nanoparticle wrapping pathway. Pedro Nunes in De Crepusculis, 1542 (Lisbon) describing the nonius, his invention Fabrice Thalmann and Carlos M. Marques Contents Bilayers and Polymers are Intimate Old Friends How do Polymers Interact with Membranes Also known as macromolecules, polymers are high molecular weight species made by the covalent binding of units called monomers. In cosmetics, pharmaceutics or detergency most formulations of membrane solutions have polymers added for performance, processing, conditioning or delivery (Lasic and Papahadjopoulos, 1998; Lasic and Barenholz, 1996; Vandepas et al. The presence of polymers in, on, or in the vicinity of a membrane changes not only the structure and viscosity of the liquid media where the membranes evolve: it modifies the properties of the membrane itself and its interactions with the environment. In this article, we introduce basic theoretical concepts for understanding, quantifying and predicting membrane transformations induced by the presence of macromolecules. We first set the stage in the next section by classifying the different modes of interaction between macromolecules and self-assembled bilayers, and by discussing the most relevant control parameters. The following sections will follow this classification, presenting polymer concepts required to the theoretical treatment of the different situations. From the point of view of the polymers, the presence of the bilayers introduces an interface, and thus an external field that will modify the state of the chains. From the point of view of the bilayer, the presence of the polymers will not only transform the membrane into a new effective membrane with different elastic properties or membrane permeability: the polymers might also induce strong shape transformations, change the structure of the membrane fluctuations and ultimately even compromise selfassembly viability. In this section, we first recall how polymers behave in the presence of interfaces and then discuss in general terms how effective elastic constants and spontaneous curvature arise for membranes in solutions containing other macromolecular species. At low enough concentrations chains do not interact, and their single chain behavior depends on the forces between monomers.
Biobran (Mgn-3). Diamox.
- How does Mgn-3 work?
- Are there safety concerns?
- Boosting immune function; preventing and treating cancer; treating AIDS, hepatitis, diabetes, or chronic fatigue syndrome; and other immune disorders.
- Dosing considerations for Mgn-3.
- What is Mgn-3?
Source: http://www.rxlist.com/script/main/art.asp?articlekey=96294
Purchase generic diamox canada
In minimally displaced or nondisplaced fractures medicine jobs purchase discount diamox on line, patients may have a painful arc of motion that is limited. Motor: Ankle dorsiflexion (deep peroneal nerve), extensor hallucis longus (deep peroneal nerve), gastrocnemius (tibial nerve), flexor hallucis longus (tibial nerve), and peroneals (superficial peroneal nerve) should be documented. Sensation: Gross sensation in sural, saphenous, deep peroneal, superficial peroneal, and tibial nerves should be documented. The quality of the pulse should be compared to the contralateral extremity and documented. If the pulses are unable to be palpated, a Doppler ultrasound should be used to document the presence or absence of pulses. In high-energy fractures with severe comminution, this step can be skipped initially due to patient discomfort. In minimally displaced or borderline operative fractures, the stability of the knee should be tested by applying a varus and valgus force with the knee in full extension. This maneuver may be too uncomfortable due to fracture pain and a large joint effusion. Any increase in joint laxity to varus or valgus stress examination greater than 10 degrees compared to the contralateral side is deemed unstable. Requires vigilant monitoring and expeditious diagnosis with subsequent surgical fascial release. The medial condyle of the tibial plateau is larger than the lateral condyle, concave, has stronger subchondral bone, and sits lower on a lateral radiograph. The lateral condyle is smaller, convex, has weaker subchondral bone, and sits higher on a lateral radiograph compared to the medial condyle. The proximal tibia articular surface has approximately 5 to 10 degrees of posterior slope (average posterior proximal tibia angle = 81 degrees). The medial and lateral menisci are fibrocartilaginous rings of tissue that are located on top of the medial and lateral condyles. These structures are frequently torn in association with tibial plateau fractures (typically reported incidence of 40% but has been described in up to 80%), most commonly tearing off its peripheral attachment to the capsule of the knee joint (meniscocapsular avulsions). These structures can be injured at their proximal or distal bony insertions while remaining attached to the bone (avulsion fracture) or can be torn in the middle of the ligament (intrasubstance tear). Patellar tendon-inserts into the anterior aspect of the proximal tibia at the tibial tubercle. Function-part of the "posterolateral corner," a group of tendons, ligaments, and the knee capsule that stabilize the knee in extension/slight flexion. Common peroneal nerve-winds around the fibular neck (upper aspect of fibula) and is the most common nerve injury in tibial plateau fractures. Deep peroneal nerve controls ankle dorsiflexion and provides sensation between the first and second web space dorsally on the foot. Superficial peroneal nerve controls ankle eversion and provides sensation to the majority of the dorsum of the foot. Course along the posterior aspect of the knee immediately behind the knee capsule. Can suffer stretch injuries due to the mechanism of injury, displaced bony fragments, or knee dislocations in association with fracture. This is due to the approximately 10-degree slope of the proximal tibia articular surface. Usually obtained for operative fractures to measure displacement and/or precisely define fracture fragments for preoperative planning. Aids the surgeon in identifying occult fractures that would otherwise be missed on plain radiographs: i. Oftentimes, the lateral meniscus is routinely visualized during surgery (see "Open reduction internal fixation"). However, during the posteromedial approach, the medial meniscus is not routinely visualized. Extra-articular (type A)-the articular surface is not fractured, but there is a fracture of the proximal tibia metaphysis that is a distance equal to or less than the width of the joint at its widest point. Partial articular (type B)-there is a fracture of the articular surface of the tibia, but a portion of the articular surface remains in continuity with the metaphysis/diaphysis. Complete articular (type C)-there is a fracture of the articular surface, and no portion of the articular surface remains attached to the underlying metaphysis/diaphysis. This classification is the most commonly used to describe the general characteristics of a tibial plateau fracture. The classification is intended to describe increasing severity of fractures with each number. Schatzker I-isolated split fracture of the lateral tibia plateau; often occurs in young patients with strong subchondral bone. Be aware of the difference between a lateral tibial plateau split fracture with displacement versus a lateral tibial plateau fracture with depression as they can be easily confused radiographically. Schatzker V-fracture of the medial and lateral condyles of the tibial plateau ("bicondylar fracture") typically with the tibial spines remaining in continuity with the diaphysis. The knee should be immobilized in order to help decrease patient discomfort with either a knee immobilizer or long leg plaster splint with the knee in slight flexion. Nondisplaced or minimally displaced lateral plateau fracture stable to stress examination. Long-term studies have demonstrated that an unstable joint significantly contributes to the development of post-traumatic osteoarthritis. The exact amount of condylar widening and joint depression that is acceptable to treat without surgery is not known. However, any joint depression or condylar widening that produces joint instability is an indication for surgery. Nonoperative treatment Avulsion fractures of the tibial plateau that do not involve a significant amount of the articular surface (typically minimally displaced tibia spine fractures). Nondisplaced medial plateau fractures are prone to late secondary displacement with nonoperative management. Indirectly reduces the fracture via fracture fragment soft-tissue attachments (ligamentotaxis). Pins are typically placed outside of the anticipated location of definitive fixation. The anterior compartment fascia and iliotibial band are incised in line with the skin incision. The anterior compartment musculature is elevated off the proximal tibia exposing the underlying anterolateral tibial plateau. A submeniscal arthrotomy may be performed at this point in time to gain access to the joint and directly visualize the joint surface. Straight incision 2 cm posterior to the posteromedial border of the tibia to the adductor tubercle of the medial femoral condyle. Alternatively, the incision can be curved 90 degrees posteriorly at the joint line. Sharp dissection through subcutaneous tissue down to fascia and pes anserine tendons. The pes anserine tendons need to be dissected to increase their mobility in order to work in the interval between the pes anserine tendons and the medial head of the gastrocnemius. Alternatively, some surgeons will elect to transect the tendons and repair them at the end of the case to enhance visualization. The gastrocnemius fascia is sharply incised along the posteromedial aspect of the tibia, exposing the underlying tibial plateau bone and fracture. The joint surface is not routinely visualized from the medial approach with intraoperative fluoroscopy used to determine reduction or arthroscopically assisted.
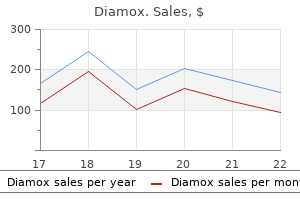
Purchase diamox 250 mg line
Complete myodesis with heavy suture to close posterior fascia to anterior periosteum; the myodesis can be completed through drill holes in the anterior tibial cortex for a more secure repair medicine ball core exercises purchase diamox online. Close the superficial subcutaneous layer with 2-0 absorbable sutures and the skin with 2-0 non-absorbable sutures, using Steri-Strips to augment the closure between sutures; then apply sterile dressings, plus/minus a removable cast. Transect anterior (quadriceps) and lateral musculature using principles described above. Identify and isolate the femoral artery and vein, suture ligate as described above. Pull traction and sharply transect the saphenous nerve allowing it to retract proximally. Transect medial musculature (adductors, sartorius, gracilis) 5 to 10 cm distal to the anticipated femoral cut for later adductor myodesis. Identify the sciatic nerve and place a single suture ligature; apply traction and sharply transect the sciatic nerve, allowing it to retract proximally. Complete the adductor myodesis with heavy suture through drill holes in the distal femoral residuum; suture the posterior hamstring fascia to the anterior quadriceps fascia for additional soft-tissue coverage of the stump. These decrease edema and mold residual limb into conical shape to accelerate prosthetic fitting. Care must be taken or it may create a tourniquet, also it needs frequent reapplication. Suture removal at 3 to 4 weeks, wound often slow to heal (particularly in dysvascular and diabetic patients). Try cortisone injections, but this often leads to revision surgery for formal neurectomy. Potential etiologies-remaining nerves continue to generate signals; spinal cord initiates excessive spontaneous firing in the absence of expected sensory input, altered signal transmission/ modulation/response within the somatosensory cortex. Home modifications-external ramps, stair lift/railings, doors widened for wheelchairs, kitchen work surfaces and sinks adjusted lower, shower with level entrance, shower seat, possible hoist for bath, adapted furniture/mirrors. Car modifications-modify pedals/seat, lift to assist entry, consider hand controls. Creation of a custom-fitted thermoplastic socket to best fit each unique residual limb. Goal is to achieve an intimate fit with a very close match between socket and residuum. Suspension system-harnesses, such as straps, belts, or sleeves, are used to attach the prosthetic limb; currently far more common for upper limb amputations. Harness is sometimes still necessary with a short residuum after a high transfemoral amputation. For some types of amputations, the prosthetic is able to stay attached by fitting around the shape of the residual limb. One of the most popular types of suspension mechanisms relies on suction-the prosthetic limb fits snugly onto the residual limb and an airtight seal keeps it in place. After preparing a preliminary socket, multiple revisions and modifications are often necessary to achieve a satisfactory fit. Much of this process is empirical, and the prosthetist makes adjustments as necessary to satisfy the user. Macroporous titanium implants fully incorporate directly into skeletal residuum, providing a solid, intimate bond between patient and prosthetic limb. Main indication is for amputees unable to use a socket-mounted prosthetic limb due to short stumps, scarred adherent skin, bony prominences, and recurrent ulcerations and infection. Concerns regarding risk of infection-when using contemporary implants and improved techniques, infection is much less problematic than anticipated. Currently no Food and Drug Administration approved implants for use in the United States, but popular internationally. Role for transhumeral amputees evolving: combined with targeted muscle reinnervation and using the most advanced myoelectric prostheses; early results are promising. K 0-unable to use/benefit from a prosthetic limb-wheelchair bound or requires crutches. K 1-uses a prosthetic limb for transfers or ambulation on a level surface-household ambulator. K 2-only able to traverse low environmental barriers, limited community ambulator. K 3-capable of variable cadence-able to traverse most environmental barriers, curbs or stairs-unlimited community ambulator. K 4-higher-demand patients, few if any restrictions including running, jumping, sports (typical of children), active adults, athletes. Morbidly obese patients-unlikely to use socket-mounted prosthesis if weigh > 150 Kg, difficult to fit socket, consider end-bearing stump (through knee, Boyd, or Pirigoff). An analysis of outcomes of reconstruction or amputation after leg-threatening injuries. A wide spectrum of injury can be encountered with rib fractures, ranging from single/nondisplaced rib fractures to multiple fractured/displaced ribs to flail chest injuries with mechanical instability of the chest wall. Increasing severity of rib injury is clearly correlated with increasing levels of morbidity and mortality. The traditional management of these injuries (including severe injuries such as flail chest) has been largely nonoperative with analgesia, supportive care of respiratory function as required, and chest tube placement for the management of associated pneumothorax/hemothorax. Recently, there has been increasing interest in the surgical management of more severe rib fractures and chest injuries. Substantial rib injuries typically occur as a result of high-energy blunt trauma (typically motor vehicle collisions, falls from 10 feet or greater, pedestrian hit by vehicle, etc. Specific physical examination of the chest should include assessment of cardiovascular function (heart rate, blood pressure, cardiac monitoring), respiratory function (tracheal deviation, oxygen saturation, air entry, percussion for dullness or hyper resonance, asymmetric or paradoxical chest movement), and physical findings of chest trauma (ecchymosis, seat-belt sign, open injuries, crepitus, subcutaneous emphysema). Commonly associated nonorthopaedic injuries include head injuries and intra-abdominal injuries such as spleen or liver lacerations. Anatomy-in addition to the anatomy of the thoracic cage and its contribution to respiratory function, it is important to recognize the anatomic structures contained within the thorax and upper abdomen by the ribs, and the associated injuries which can occur to these structures. Mediastinal structures including the heart (cardiac tamponade, cardiac contusion, laceration), great vessels (vascular injury), trachea (tracheobronchial injury), and esophagus (esophageal rupture). Pleural space (tension pneumothorax, sucking chest wound, pneumothorax, and hemothorax). Ribs and thoracic cage (increasing severity of rib injuries cause increasing impairments in ventilatory function due to both painful inspiration/expiration and compromised respiratory mechanics). He was transferred to our Level 1 trauma center with multiple suspected right-sided rib fractures after being intubated. His chest X-ray shows a large pneumothorax (blue arrows) and tracheal deviation (red arrow) consistent with a tension pneumothorax. Serves as the initial screening test for the identification of rib fractures, pneumo/hemothorax, and diaphragmatic rupture. Although no widely recognized classification system exists for rib fractures or bony injuries to the thoracic cage, it is important to recognize that a wide spectrum of pathology exists. Muscle-splitting windows in the serratus anterior (white arrow) and external oblique (blue arrow) have been used to access rib fractures. The literature has clearly shown that as the severity of these injuries increases, there are substantial increases in both morbidity and mortality. Flail chest injuries occur when multiple ribs have segmental fractures, creating a "flail segment" which moves paradoxically with respiration (inward during inspiration and outward during respiration). Three or more unilateral rib fractures combined with sternal fracture or dissociation. Initial management is often directed at the treatment of associated injuries and supporting ventilation with the goals of maintaining oxygenation and controlling hemorrhage. Both the initial and definitive treatment of patients with severe rib injuries often requires multidisciplinary assessment and treatment involving orthopaedic surgery, general surgery or thoracic surgery, and intensive care specialists.
Order diamox amex
These observations can be understood from the competition of different energy contributions which favor necklace-like tubes below a certain critical tube length but cylindrical tubes above this length (Lipowsky medications like tramadol order diamox paypal, 2013; Liu et al. At the critical tube length, the necklace-like tube transforms into a cylindrical one. The existence of a critical tube length can be understood intuitively from the following simple argument (Lipowsky, 2013). If the membrane has spontaneous curvature m, a necklace-like tube consisting of spherules with radius R 2 = 1/ m connected by closed membrane necks has vanishing bending energy. For a cylindrical tube with radius Rcy = 1/(2 m), the main body of the cylinder also has vanishing bending energy but such a tube must be closed by two end caps which have the finite bending energy 2. On the other hand, the necklacelike tube has a larger volume compared to the cylindrical one and the osmotic pressure difference across the membranes acts to compress the tubes when they protrude into the interior solution within the vesicles. Therefore, such a tube can lower its free energy by reducing its volume which favors the cylindrical tube. The volume work is proportional to the tube length whereas the bending energy of the end caps is independent of this length. The competition between these two energies then implies that short tubes are necklace-like whereas long tubes are cylindrical. The same conclusion is obtained by minimizing the bending energy of the whole vesicle membrane (Liu et al. One then finds that, for fixed vesicle volume and membrane area, the mother vesicle has a smaller bending energy when it forms a cylindrical tube and that this energy decrease of the mother vesicle overcompensates the bending energy increase from the end caps of the cylinder when the tube is sufficiently long. The critical tube length at which the necklace-like tube transforms into a cylindrical one is about three times the vesicle radius. For large vesicles, the adhesion energy must dominate because it is proportional to the contact area of the vesicle and thus grows quadratically with the size of the vesicle whereas the increased bending energy is concentrated 5. We are then left with only three dimensionful parameters, the membrane area A, the bending rigidity, and the adhesive strength W. The non-adhering or free vesicle forms a spherical shape Sfr with bending energy beSfr = 8. When the vesicle membrane spreads onto an adhesive surface, the vesicle attains the shape Sad with contact area Abo of the bound membrane segment and gains the adhesion energy Ead - W Abo. In order to study the overall shape of the adhering vesicle, one may ignore the molecular details and focus on the adhesive strength W of the membrane-surface interactions which corresponds to the adhesion (free) energy per area (Seifert and Lipowsky, 1990). This coarse-grained description of the membrane-surface interactions in terms of the single parameter W is consistent with the separation of length scales that has been used to construct the different curvature models. Because the bound and the unbound membrane segments are exposed to different environments and, thus, to different molecular interactions, they can differ in their molecular composition and, thus, in their curvature-elastic properties (Rouhiparkouhi et al. In order to reduce the number of parameters, we will first assume that this ambience-induced segmentation of the vesicle membranes can be ignored and that the bound and unbound membrane segments have the same curvature-elastic properties. Adhesion-induced segmentation of multi-component membranes will be discussed at the end of this section and at the end of Section 5. Furthermore, we will again focus on the spontaneous curvature model which depends on only two dimensionless parameters, the volume v and the spontaneous curvature m. When we parametrize the adhesion energy in terms of the dimensionless adhesive strength w proportional to W /, vesicles adhering to planar surfaces are described by only three parameters. On the one hand, this parametrization is convenient from a theoretical point of view because it allows us to explore large regions of the parameter space. On the other hand, the additional parameter W can be directly deduced from experimental observations of adhering vesicles. At the end of this section, more complex adhesion geometries will be briefly discussed corresponding to curved and/or chemically patterned substrate surfaces. The extension of the theory described here to the interactions of membranes with adhesive nanoparticles is described in Chapter 8 of this book. For a planar surface, this adhesion energy is the only energy contribution from the bound membrane segment. The unbound membrane segment, on the other hand, has to adapt its shape to the presence of the substrate surface which leads to the bending energy increase Ebe = beSad - beSfr = 8 Ebe. In general, the adhesion of vesicles involves three additional parameters: the osmotic conditions that determine the volume-toarea ratio, the spontaneous curvature m of asymmetric bilayers, and the mean curvature Mbo of the bound membrane segment arising from a curved adhesive surface. In order to take these additional parameters into account, we need a systematic theory based on an appropriate energy functional. The total membrane area A can then be decomposed according to A = Abo + Aun = Sbo + Sun (5. In general, the two partial areas also depend on the shape of the adhesive surface. In other words, the membrane shape should not exhibit any kink along the contact line. This geometric requirement is equivalent to the condition that the membrane has a finite bending energy (Seifert and Lipowsky, 1990). Because the normal vector is required to vary continuously across the contact line, the principal curvature C co tangential to the contact line vanishes. In addition, the principal curvature Cco of the unbound membrane segment perpendicular to the contact line is given by C co = 2 W / (5. Therefore, the contact mean curvature becomes 1 1 M co = (Cco + C co) = C co = W /(2) (planar substrate). It is interesting to note that the contact mean curvature Mco does not depend on the spontaneous curvature m, which is somewhat counterintuitive. This m-independence also applies when the vesicle adheres to a curved surface, see further below. However, the shape and the contact area of an adhering vesicle do depend quite significantly on the spontaneous curvature (AgudoCanalejo and Lipowsky, in preparation). One should also note that the principal curvature C co jumps along the contact line from C co = 0 within the bound membrane segment to C co = 2 W within the unbound / segment. Likewise, as mentioned, the mean curvature jumps from M = 0 within the bound membrane segment to M = M co within the unbound segment. In the following sections, we will see that analogous curvature discontinuities are also present along domain boundaries separating two intramembrane domains and along three phase contact lines arising from membrane wetting. Adhesion length with the constraints that S = V and S = A where V and A are the prescribed vesicle volume and membrane area as before. It is important to note that the value of the contact area Abo of the bound membrane segment is not prescribed here which implies that the contact line is not pinned but free to find its optimal position. Additional parameters related to adhesion As before, it is again convenient to choose the vesicle size Rve = A /(4) as the basic length scale and the bending rigidity as the basic energy scale. The shape of the adhering vesicle then depends on the dimensionless volume v = 6 V / A 3/2 and on the dimensionless spontaneous curvature m = m Rve, both of which also determine the shape of free vesicles. In addition, the adhering shape also depends on the dimensionless adhesion strength 2 w W Rve / (5. The simplest substrate geometry is provided by a planar surface with Mbo = 0 which reduces the parameter space to the three dimensionless parameters v, m, and w. The next-to-simplest substrate geometry is obtained for constant-mean-curvature surfaces such as spherical surfaces or cavities. In the latter case, the mean curvature M bo of the bound membrane segment is constant and the parameter space becomes four-dimensional. In the following subsections, we will first discuss the planar case and subsequently summarize the modifications arising from spherical surfaces and cavities. We require the bound and the unbound membrane segments to join along the contact mean curvature M co = W /(2) as given by Eq 5. When the adhesion length becomes of the order of 10 nanometer as in the first two rows of Table 5. In all panels of this figure, the membrane has the same area and the same bending rigidity as well as vanishing spontaneous curvature. The five vesicle shapes are obtained for five different values of the adhesive strength w. Thus, the vesicle starts to spread over the substrate surface provided (Seifert and Lipowsky, 1990) 2 w = W Rve / > w ad = 2 for P = 0. The latter criterion directly reflects the competition between the adhesive strength W and the bending rigidity which favors and disfavors the onset of spreading, respectively. Because a sphere with P = 0 is stable for m < 3/Rve, the threshold value wad = 2 for the onset of spreading is expected to apply for this range of m-values as well. This expectation is confirmed by numerical energy minimization for axisymmetric shapes (Agudo-Canalejo and Lipowsky, in preparation). The latter calculations also show that the contact area increases with increasing spontaneous curvature m > 0 even though the contact mean curvature Mco does not depend on m.
Order diamox online pills
One-sample (paired) t test or Wilcoxon rank sum test-tests the likelihood that two different measurements in the same sample are different medicine to induce labor buy diamox with a mastercard. Multiple regression-describes a numerical relationship between one dependent variable and multiple (at least two) other covariates. Careful review of inclusion/exclusion criteria and interventions (and control groups). Selection bias-occurs when the sample analyzed is not representative of the intended population. Problematic with retrospective trials as treatments provided may have been selected based upon surgeon preference, for example, unblinding of interventions in randomized trials. Are the results presented in abstract consistent with results presented in body of manuscript Carefully pay attention to figures and tables (some journals require that all results be presented in table and figure form in addition to prose). Often set the context for the research question and can provide context for use of the results in the scheme of current practice. A reference to a textbook or review article which misquotes the literature is misleading (perhaps unintentionally). Blackwell Science Ltd, Oxford, United Kingdom; 2003 39 6 Acute Infection Following Musculoskeletal Surgery Frank R. Obremskey Introduction Postoperative infection following internal fixation involves the soft tissues (skin, subcutaneous tissues, muscle fascia, and muscle), hardware, and potentially the bone. Purulent discharge from the surgical site and/or incision with or without associated erythema, tenderness, or fever. Symptoms (local or regional pain or joint stiffness) which may be less obvious signs of infection. Absence of radiologic evidence of bone healing after several months, with or without fixation failure, may also suggest infection. Intermittent fevers, chills, sweats (particularly, night sweats in the setting of chronic infections), and general malaise are common symptoms. An untreated infection may progress rapidly and threaten the limb, lead to septic shock, or even lead to death. High suspicion is necessary for post-operative surgical sites with atypical findings or patient reporting increased pain. Affects the incision but does not extend to the fracture site and remains superficial to the level of the fascia. Surgical devices represent a substrate for microbial colonization and biofilm-associated infection. Variety of organisms have been associated with indwelling implants, some of the most common are: i. Planktonic bacteria attachment: reversible and bacteria susceptible to antibiotics and rinsing. Micro-colonies develop: reversible and bacteria susceptible to antibiotics and rinsing. Continued cell division: more adhesion sites, matrix formation, and biofilm maturation. Detachment: liberate planktonic bacteria or small segments and plankontic bacterial may relocate and colonize other surfaces. Persister cells: dormant, multidrug tolerant cells that live within mature biofilm and have the ability to repopulate the biofilm. Quorum-sensing molecules: chemomodulators within a mature biofilm permitting intercellular communication to permit bacterial resistance. Twenty percent of patients undergoing nonunion repair with normal preoperative inflammatory markers may be culture-positive at the time of surgery. Diagnostic imaging in the weeks immediately following operative care often fails to show changes that are commonly seen over the course of time. Computed tomography or ultrasound may provide findings of an abscess or presence of air. Such findings may either guide percutaneous drainage with a needle or direct surgical debridement. Classification Infections are typically referred to as superficial or deep according to whether the infection has penetrated deep to the fascia. Retention versus removal of implants with staged internal fixation after temporary fixation (typically external fixation). Modifiable risk factors should be addressed to optimize treatment(s) as local host factors related to reduced host vascularity, neuropathy, trauma, and immunodeficiency increase the likelihood of infection. Predictors of treatment failure include: Biopsy 43 General Principles of Orthopaedic Trauma G. If implants are removed prior to fracture healing, ensure that fracture stabilization is achieved. Implant retention-success rates of curing early postoperative infection with maintenance of hardware range from 68 to 90% with surgical debridement and treatment with culture-specific antibiotics. Implant removal-successful eradication of infection reaches 92% before bony union. Recurrence of infection following successful bony healing requires removal of hardware, debridement, and treatment with antibiotics. By following standardized diagnosis and treatment regimens outcomes can be optimized. Surgeons need to assure diagnosis of infection, optimize the patient by improving host factors as much as possible and utilizing a multidisciplinary team. The surgeon then needs to decide to retain or remove implants with a immediate or staged revision fixation. Antibiotics should be culture driven if possible and can be administered intravenous or by oral methods. Without a standardized process and multidisciplinary team patients are at risk for persistent infection and/or amputation. Maintenance of hardware after early postoperative infection following fracture internal fixation. Outcome of penicillin-susceptible streptococcal prosthetic joint infection treated with debridement and retention of the prosthesis. Role of rifampin for treatment of orthopedic implant-related staphylococcal infections: a randomized controlled trial. Hadeed Introduction the goal of orthopaedic fracture care is to treat fractures in a way that minimizes complications while maximizing functional outcomes. It is critical to understand both the natural history and effect of interventions on bone healing as operative indications are often based on the ability to decrease the chance of nonunion and malunion. When a patient develops a nonunion or a malunion, the cost to the health care system and society is great, as it typically results in multiple surgical procedures and extended time away from normal activities. A tibial nonunion has been compared to having an effect on health and wellbeing similar to some cancer or other chronic illness diagnoses. To understand malunions and nonunions, it is critical to have a basic understanding of bone healing and the biomechanics of fracture repair (discussed in depth in Chapter 1, Physiology of Fracture Healing, and Chapter 4, Biomechanics of Internal Fracture Fixation). When approaching these difficult cases, it is important to have a stepwise, reproducible approach, make the diagnosis using the history, physical exam, laboratory and radiographic data. Based on patient-specific variables, develop a treatment plan with a reasonable chance of success (Video 7. Assessment of Nonunions Factors leading to nonunion can generally be grouped into two categories: biologic and mechanical. The assessment is a gathering of data on known factors which may have contributed to a failure of the biologic and mechanical success of the fracture healing. Symptoms (or history of symptoms) associated with infection: erythema, swelling, drainage, fevers, chills. The data of patient-specific risk factors is obtained after completing a thorough history with each individual patient. Nonsteroidal anti-inflammatory drugs negatively affect the pathways responsible for bone healing. In some studies, female patients and older patients had an increased rate of nonunion. Diseases that negatively affect vascularity, such as diabetes mellitus and other vascular disorders, can impair fracture healing.
Order diamox 250mg without prescription
This drug has been loaded into the bilayer core of polymersome membranes by injecting a solution of the drug (in methanol) into the polymersome suspension treatment emergent adverse event cheap 250mg diamox. Using this procedure, efficiencies higher than those for liposomes (Ahmed and Discher, 2004) have been obtained. The membrane can also be utilized to incorporate fluorescent dyes for imaging and tracking in vivo (Lin et al. Delivery of two such drugs acting via separate cytotoxic pathways might reduce incidence of drug resistance by cancer cells, and is thus one of the most promising applications of polymersomes. Polymersomes loaded with both Paclitaxel and Doxorubicin led to tumor shrinkage in vivo (Ahmed et al. The maximum tolerated dose of drugs was several-fold increased by loading in polymersomes (compared with free drug), with unloaded polymersomes themselves being benign (Ahmed et al. In nude mice implanted with human derived tumor xenografts, intravenous injections of Doxorubicin and Paclitaxel encapsulated within polymersomes led to higher drug fluorescence in tumors, while producing consistent shrinkage. Dual loaded polymersomes also led to enhanced tumor cell apoptosis compared with free drug, with levels remaining steady with time (Ahmed et al. However, the presence of active targeting groups on the surface can also reduce circulation time (Discher et al. Another important application of polymersomes has been as microreactors (Price et al. Multicompartmentalized microreactors have been synthesized to enclose different conditions in different compartments (Chandrawati and Caruso, 2012; Elani et al. Since their conception, polymersomes have served as an important tool for the field of membrane physics while also being developed for a wide range of applications. Self-assembled block copolymer aggregates: From micelles to vesicles and their biological applications. Chandrawati R, and Caruso F (2012) Biomimetic liposome-and polymersome-based multicompartmentalized assemblies. Chidambaram M, Manavalan R, Kathiresan K (2011) Nanotherapeutics to overcome conventional cancer chemotherapy limitations. Choucair A, Lim Soo P, Eisenberg A (2005) Active loading and tunable release of doxorubicin from block copolymer vesicles. Graff A, Sauer M, Van Gelder P, Meier W (2002) Virus-assisted loading of polymer nanocontainer. Kazunori K, Masayuki Y, Teruo O, Yasuhisa S (1993) Block copolymer micelles as vehicles for drug delivery. Kita-Tokarczyk K, Grumelard J, Haefele T, Meier W (2005) Block copolymer vesicles-Using concepts from polymer chemistry to mimic biomembranes. LoPresti C, Lomas H, Massignani M, Smart T, Battaglia G (2009) Polymersomes: Nature inspired nanometer sized compartments. Meng F, Zhong Z, Feijen J (2009) Stimuli-responsive polymersomes for programmed drug delivery. Messager L, Gaitzsch J, Chierico L, Battaglia G (2014) Novel aspects of encapsulation and delivery using polymersomes. Nuss H, Chevallard C, Guenoun P, Malloggi F (2012) Microfluidic trap-and-release system for lab-on-a-chip-based studies on giant vesicles. Pata V, and Dan N (2003) the effect of chain length on protein solubilization in polymer-based vesicles (polymersomes). Picker A, Nuss H, Guenoun P, Chevallard C (2011) Polymer vesicles as microreactors for bioinspired calcium carbonate precipitation. Self-assembly and properties of diblock copolymers by coarse-grain molecular dynamics. Key roles for chain flexibility in block copolymer membranes that contain pores or make tubes. Coarse-grain molecular dynamics simulations of diblock copolymer surfactants interacting with a lipid bilayer. Walde P, Cosentino K, Engel H, Stano P (2010) Giant vesicles: preparations and applications. Ludwig Erhard Contents Introduction Criteria to Be Fulfilled to Obtain Hybrid Giant Vesicles 27. Their major characteristics are described and discussed in Chapter 26, to be consulted by readers who are non-specialist in the field before going through the present chapter. This one deals with hybrid, that is, intimately mixed polymer/lipid vesicles that can be viewed as advanced vesicular structures as compared with their liposome and polymersome forerunners, as they potentially marry in a single membrane the best characteristics of the two separate components. Ideally, these structures could present biocompatibility and bio-functionality of liposomes, as well as robustness, low permeability and functional variability conferred by the copolymer chains. This should be of great interest in pharmaceutical applications for which only a few formulations based on liposomes are commercially available despite decades of research. In particular such moderate use of liposomes in clinics could be due to their lack of mechanical stability in the high shear rate of blood circulation through tiny vessels. As a consequence, the controlled release of encapsulated molecules at the predetermined biological target. Promising results have been obtained regarding their drug targeting ability and biomolecular recognition properties (Cheng et al. To date, the physical and molecular factors governing the phase separation in these hybrid copolymer/lipid membranes are only 552 Giant hybrid polymer/lipid vesicles partially understood. In addition to the expected chemical incompatibility between copolymer block chains and phospholipids, one also has to consider the respective dimensions of the molecules as well as those of the corresponding bilayers. In order to perfectly benefit from the potential of such systems, the membrane structure must be tuned either toward homogeneous mixing of the molecular components or, on the contrary, toward lateral phase separation leading to the presence of nano/micrometric domains. The relationship between membrane structure and physical and bio-functional properties must then be better understood in order to eventually optimize and validate the use of hybrid vesicles in future applications like drug delivery, tumor targeting, bio-recognition or bio-adhesion. It has to be noted that hybrid vesicles reported over the last 10 years were exclusively prepared by a one-step process by which a film composed of the desired amount of copolymer and phospholipid is hydrated. We will not describe previous approaches based on the modulation of lipid membrane properties by adsorption of amphiphilic polymers onto preformed giant liposomes. These systems have been proposed as tools to understand the structure-properties relationship of biological cell membranes (both of the plasma membrane and that of the internal organelles), which are constituted of different lipids and membrane proteins ensuring part of the many biological functions of the cell: transport of matter, energy, cell division, signaling pathways. The lipid composition in the membrane strongly depends on the nature of the cell (eukaryote, prokaryote or archaea) and comprises several classes: glycerophospholipids, sphyngolipids, sterols, saccharolipids, and so on. This subtle association allows flexibility and fluidity of the membrane and the formation of lipid raft like domains, which can arise from the aggregation of proteins such as clathrin but can also be driven by lipid segregation. Lipid rafts are mainly composed of sphingolipid and cholesterol-rich domains and contain a variety of signaling proteins. It has been established that the lipid rafts play an important role in health and disease (Michel and Bakovic, 2007). Basically, two types of phase separation in lipid membrane can occur: Lateral phase separation of two lipids into different areas or orthogonal phase separation between the two leaflets of the lipid bilayer. Orthogonal phase separation can be triggered via addition of an external compound. Nonideal mixing or even demixing (phase separation) can occur between lipids with similar structure. In that case, phase separation is obtained below a given temperature of fluid/solid or solid/solid transition. A strong difference in melting temperatures is generally associated with a strong difference in chemical structures. Cholesterol has been largely employed to modulate the fluidity of membranes and to create phase separation above the main transition temperature of a phosphocholine lipid. Phase separation leads to lipid/lipid boundaries and possibly to a height mismatch between both phases. Consequently, the membrane elastically deforms at the domain interface to minimize the exposure of hydrophobic tails to water. The height mismatch has an energetic cost proportional to the length of the boundary line, thus defining the line tension. Thermodynamically, the line tension tends to favor domain coalescence (once a nucleation size is reached) to minimize the boundary length. As a consequence, fluid lipid domains should grow with time into one single large-circular domain in the membrane.
Syndromes
- Another class of hormone therapy medicines is called aromatase inhibitors. Medicines such as exemestane (Aromasin) work as well or even better than tamoxifen in postmenopausal women with breast cancer. Aromatase inhibitors block estrogen from being made in the body.
- Hematoma (blood accumulating under the skin)
- Drowsiness
- Bleeding disorders
- The skin or mouth lesions change in appearance
- Irritable bladder
- Kidney damage due to the contrast dye (more common in patients with diabetes or kidney problems)
- Paints
- Difficulty lifting up the foot and toes and making toe-out movements
- Paromomycin
Generic 250mg diamox with visa
Svetina S medicine abuse buy 250mg diamox with amex, Zeks B (1989) Membrane bending energy and shape determination of phospholipid vesicles and red blood cells. Veatch S, Keller S (2003) Separation of liquid phases in giant vesicles of ternary mixtures of phospholipids and cholesterol. Walde P, Cosentino K, Engel H, Stano P (2010) Giant vesicles: Preparations and applications. Giant vesicles theoretically and in silico 6 Simulating membranes, vesicles, and cells the form is the outer expression of the inner content. Fedosov, and Gerhard Gompper Contents Introduction Membrane Models and Simulation Techniques 6. A biomembrane is typically composed of many different lipids, which gives a cell many opportunities to control membrane properties by adjusting the membrane composition. This can modify the spontaneous curvature and the bending rigidity, and even lead to phase separation and domain formation. In addition, a biological membrane contains a large number of trans-membrane proteins, which control the exchange of water, ions, and small molecules between the cell plasma and the extracellular space. Vesicles are cells striped down to the minimum, a membrane enclosing a fluid volume. Vesicles are therefore ideal model systems to investigate the physical properties of many components of cells in isolation, without the full complexity of the cellular machinery. Because the systems are well defined, their properties can be analyzed and studied much more easily from a theoretical perspective. Although vesicles are relatively simple, they are still complex many-body systems, with lipids forming a bilayer due to the hydrophobic effect, a plethora of shapes and phase transformations, families of different genus, membranes with holes, and so on. Simulations therefore play a very important role in elucidating their equilibrium and dynamic properties. Here, simulation approaches range from the molecular scale-where the properties of lipids and membrane proteins are studied-over the supramolecular scale-where the self-assembly of lipids and their phase-behavior can be investigated-to the vesicle scale-where shapes and shape transitions, the effect of phase separation in the membrane and the internal fluid, and the deformations due 170 Simulating membranes, vesicles, and cells Giant vesicles theoretically and in silico to external forces and fluid flow are studied (see also Chapters 7, 15, and 19). Simulations are also important because the focus of the research is shifting from simple single-component to biologically more relevant multicomponent systems. Therefore, several different models, which are suitable to study phenomena on a smaller range of length scales as illustrated in Box 6. The interactions are sometimes treated quantum-mechanically, but are modeled in most cases using classical force fields. All-atom simulations are indispensable whenever the chemical structure of the participant molecules is relevant for the phenomena under investigation. For example, the functioning of membrane proteins that act as ion pumps can only be understood on the basis of such atomistic models. In such a model, water becomes a Lennard-Jones fluid with attractive interactions, and amphiphilic molecules become short polymer chains with two kinds of monomers, with attractive or repulsive interactions with the solvent particles and the other monomers (den Otter and Briels, 2003; Goetz et al. Very similar models, with Lennard-Jones interactions replaced by linear "soft" potentials, have also been employed intensively in dissipative particle Box 6. Solvent-free bilayer model (Noguchi and Takasu, 2001a, 2001b); reprinted from Gompper and Noguchi (2006). Triangulated surface model (Gompper and Kroll, 1997, 2004); reprinted from Gompper and Noguchi (2006). Meshless membrane model; adapted with permission from Noguchi and Gompper (2006b). Different models and simulation techniques are required to capture the behavior at different scales. The coarse-grained modeling can be taken one step further by taking into account the different chemical nature (and electrical charge) of various head- and tail-groups (Marrink and Mark, 2003; Marrink et al. This allows progress from a more qualitative to a more quantitative description of membrane properties. Such models allow molecular dynamics simulations of few thousand lipids and make it possible to study the formation, structure, and dynamics of small phospholipid vesicles (Marrink et al. Solvent-Free Membrane Models-The solvent in a coarsegrained model is required for two reasons. First, it is necessary to stabilize the bilayer structure due to the repulsion between the solvent and the amphiphile tails. Second, it mediates hydrodynamic interactions between different parts of the membrane. However, the simulation of the motion of solvent particles consumes a large fraction of the total simulation time. Therefore, solvent-free membrane models have been designed that work as well as the models with solvent when structural and thermodynamic properties are investigated. Additional interactions between amphiphiles have to be introduced in this case in order to mimic the hydrophobic interactions with the solvent (Brannigan and Brown, 2004; Cooke and Deserno, 2005; Cooke et al. This approach is advantageous in the case of membranes in dilute solution because it reduces the number of molecules by orders of magnitude. However, the basic length scale is still on the order of magnitude of the size of the amphiphilic molecules. Triangulated Surface Models-The natural length scale of the previous two classes of membrane models is the size of the head group of a lipid molecule, that is, roughly 1 nm. In this case, a continuum description on the level of elasticity theory is required. In order to make such continuum models amenable to computer simulations, triangulated surfaces are often employed (Gompper and Kroll, 1997, 2004). The building blocks in such models correspond to membrane patches consisting of hundreds or thousands of lipid molecules. The main idea here is to connect membrane "nodes" (or "vertices") by a triangular network of bonds. The bond potentials are chosen so as to achieve a homogeneous distribution of vertices on the membrane. For fluid membranes, this requires a dynamic triangulation such that vertices can diffuse and flow within the membrane. For polymerized membranes, such as in capsules, a fixed connectivity represents the unbreakable bonding between neighboring molecules, and implies a shear elasticity of the membrane. Meshless Membrane Models-A different approach to discretize elasticity theory of a two-dimensional (2D) surface embedded in 3D space is to employ an ensemble of membrane nodes without connecting them to form a triangulated mesh. Meshless membrane models instead employ pairwise and multi-particle interactions to (i) achieve a roughly homogeneous density of nodes on the membrane and (ii) favor smoothly curved membrane conformations (Noguchi and Gompper, 2006b). The advantage of meshless membrane models is that open boundaries-which occur, for example, in membrane rupture-and topology changes-as in vesicle fusion-can be very easily simulated. The quality of the results depends crucially on the development of reliable force fields that allow a quantitative description of the collective behavior of many molecules. For lipids, experimental reference data are employed that include specific structural properties of lipid bilayers, such as area per lipid, volume per lipid, bilayer thickness, order parameter for the lipid tail orientation, and headgroup hydration. Several such force fields have been developed and tested in recent years (Dickson et al. Not only all-atomistic, but also some coarse-grained force fields retain chemical specificity. After equilibration, the system can be transformed (back) to a relaxed atomistic model if required. In particular for complex multicomponent membranes, a chemically specific coarse-grained approach provides an efficient means for generating equilibrated atomistic models. However, the results on the coarse-grained level can also be interpreted directly without a subsequent atomistic simulation. It emphasizes the universal aspects that are common to many different amphiphilic systems, independent of the detailed chemistry of a particular system. In this case, the Lennard-Jones interactions between two particle species i and j are replaced by the conservative forces FijC = a ij (1 - rij /r0) rij (6. The amphiphilic molecules are modeled as short polymeric chains with head (H) and tail (T) particles, so that neighboring particles in the chain interact via the harmonic-spring potential U chain (r) = kchain (r -)2. Head particles mutually attract each other with the Lennard-Jones potential in Eq.
Diamox 250mg otc
When we deflate such a two-droplet vesicle treatment associates diamox 250 mg without a prescription, it can decrease its interfacial energy by reducing the area A of the interface. The corresponding energy gain is governed by A where A is the change in interfacial area. Such a morphological change is, in fact, rather likely unless one of the membrane segments has a sufficiently large spontaneous curvature to form nanobuds and nanotubes. If the segment forms nanotubes, for example, the energy gain is A with the area A stored in the nanotubes 2 and the spontaneous tension = 2 m. So, we expect that osmotic deflation of a partially wetted vesicle leads to a reduction of the interfacial area whenever. This competition between different morphological pathways is more systematically described in Appendix 5. Therefore, we obtain the stability criterion E1 0 which is equivalent to 1 (M - m) + (M - m) co (G, G,). Because the additional term arising from the interface is irrelevant in the limit of small neck radius R ne, 144 Understanding giant vesicles: A theoretical perspective the stability criterion in Eq. The latter assumption is justified for a positive value of the contact line tension co but may not apply to a negative value of co. Indeed, recent molecular simulations have shown that a negative contact line tension can lead to a spontaneous symmetry breaking of the rotational symmetry and to a tight-lipped contact line (Satarifard et al. In the nucleation regime, the droplets are formed by the minority phase and have to overcome a certain free energy barrier in order to grow. For a rigid surface as provided by a tense membrane, the barrier reduction depends primarily on the contact angle of the droplet. For a flexible and deformable membrane, as considered here, the barrier may be further reduced by the elastic response of the membrane which can adapt its shape and composition to the molecular interactions with the droplet. As in the previous subsections, we focus on phase separation of the interior aqueous solution into two coexisting liquid phases, and. Both segments are exposed to asymmetric aqueous environments which act to induce spontaneous curvatures m and m. Here, we focus on the case m 0 and m m: (a) Initially, the interface has the shape of a spherical cap and forms the * intrinsic contact angle with the adjacent segment (blue) of the membrane; (b) For negative values of m, the membrane segment prefers to form a spherical in-bud that is filled with exterior phase. For such a small droplet, the intrinsic contact angle will be affected by the tension co of the contact line, see Eqs 5. This contact line tension can be positive or negative, in contrast to the line tension of domain boundaries which is always positive. In fact, recent molecular simulation indicate that the contact line tension co can be negative (Satarifard et al. In order to simplify the following discussion, let us assume that the spontaneous curvature m is large compared to the spontaneous curvature m of the segment and that the latter curvature is small and can be ignored. As shown in this figure, all membrane segments adjacent to the closed neck are formed by the membrane with spontaneous curvature m. The membrane neck is then characterized by the condition 1 0 > M ne = (M1 + M 2) m where M1 and M2 = -1/R are 2 the mean curvatures of the two membrane segments 1 and 2 on the two sides of the neck. Because these two membrane segments have the same curvature-elastic properties, this stability condition is identical with Eq. As shown in the latter panel, the two membrane segments adjacent to the neck of the out-bud are now provided by the and the segments which have, in general, different spontaneous curvatures m and m. The formation of the out-bud reduces the free energy of the membrane-droplet system by (i) adapting the mean curvature of the segment to its spontaneous curvature m and (ii) replacing the interface by a closed membrane neck which implies a strong reduction of the interfacial free energy. Now, let us briefly consider two important topology-transforming processes, membrane fusion and membrane fission (or scission). During membrane fusion, two separate membranes are combined into a single one; during fission, a single membrane is divided up into two separate ones. These processes are ubiquitous in eukaryotic cells: Both the outer cell membrane and the inner membranes of organelles act (i) as donor membranes that continuously produce vesicles via budding and fission and (ii) as acceptor membranes that integrate such vesicles via adhesion and fusion. One example for fission is provided by the closure of autophagosomes which are double-membrane organelles (Knorr et al. Fusion is exergonic, if the free energy G2 of the 2-vesicle state exceeds the free energy G1 of the 1-vesicle state. Dependence on spontaneous curvature the free energy difference G2 - G1 between the 2-vesicle and the 1-vesicle state can be estimated if one ignores energetic contributions arising from changes in volume and focuses on changes in curvature energy (Lipowsky, 2013). Because of the topological changes, we need to take the Gaussian curvature and the associated Gaussian curvature modulus G into account. For the following considerations, it will be sufficient to use the rough estimate G - which is consistent with both experimental (Derzhanski et al. A small spherical vesicle that is cleaved off from a donor membrane then changes the total curvature energy by a certain amount that can be used to estimate the free energy difference G2 - G1. It is important to note, however, that this change in curvature energy depends strongly on the magnitude of the spontaneous curvature. We may then ignore any constraints on the vesicle volumes and assume that the large vesicle of the 2-vesicle state has a spherical shape as well. If this bud is cleaved off, the free energy difference between the resulting 2-vesicle state and the initial 1-vesicle state is now negative and given by G2 - G1 = 8 (1 - 2R ss m) + 4G 4G -4 for Rss 1/(2 m). In this case, the free energy G2 of the 2-vesicle state exceeds the free energy G1 of the 1-vesicle state; and (b) Schematic landscape for an exergonic fission process. In the latter case the free energy G1 of the 1-vesicle state is larger than the free energy G2 of the 2-vesicle state. In both (a) and (b), the free energy difference G2 - G1 determines the direction in which the processes can proceed spontaneously (black arrows) while the kinetics of these processes is governed by the free energy barriers. Biological membranes often form intramembrane domains with an appreciable spontaneous curvature mdo. One example for this latter case is provided by clathrin-dependent endocytosis which leads to membrane domains with a spontaneous curvature mdo -1/(40nm) (Agudo-Canalejo and Lipowsky, 2015a). The membrane domain can then form a small spherical bud of size R ss = 1/ mdo as follows from the closed neck condition for domain-induced budding, see Eq. If the latter bud is cleaved off, the free energy difference between the resulting 2-vesicle state and the initial 1-vesicle state is again negative and has the form G2 - G1 = 8 (1 - 2Rss mdo) + 4G - 4 - 12 - 4 mdo mdo (5. Because the vesicle membranes are fluid, they can respond to external perturbation by remodeling both their shape and their local membrane composition. Two curvature-elastic parameters that play a prominent role in the whole chapter are the spontaneous curvature m, which provides a quantitative measure for bilayer asymmetry (Section 3. If molecular flip-flops between the two leaflets of the bilayer membrane can be ignored, the spontaneous curvature becomes an effective spontaneous curvature meff that contains both a local and a non-local contribution, the latter arising from area-differenceelasticity, see Eqs 5. All biomembranes are asymmetric in the sense that the two leaflets have different lipid compositions (Fadeel and Xue, 2009) and that the membrane proteins have a preferred orientation related to their biological function. It is important to realize that both lipids and membrane proteins as well as adsorbed solutes and anchored macromolecules can contribute to the spontaneous curvature as illustrated by the examples in Box 5. In fact, the framework of curvature elasticity as reviewed here applies to giant vesicles irrespective of the chemical nature of the molecular membrane components as long as the vesicle membranes are in a fluid state. Thus, these vesicles may be built up from different lipid components, membrane proteins, or other amphiphilic molecules such as diblock copolymers. The shapes and shape transformations of membranes with laterally uniform curvature-elastic properties are governed by two dimensionless parameters, the volume-to-area ratio (or reduced volume) v and the spontaneous curvature m = Rve m. These two parameters can be controlled by changes in the osmotic conditions and by one of the curvature-generating mechanisms in Box 5. The resulting shape transformations often lead to budding and tubulation processes, which create nanobuds and nanotubes as described in Sections 5. The buds and tubes represent additional membrane compartments that are still connected to the mother vesicle via closed or narrow membrane necks. The latter deduction is based on the local stability conditions for closed necks as given by Eqs 5. In the absence of flip-flops, one obtains the generalized stability conditions in Eqs 5. Sufficiently large values of meff lead to the cleavage of the membrane neck and thus to complete membrane fission, see Section 5. In cell biology, the closure and cleavage of such membrane necks represents an essential step for many processes such as endo- and exocytosis, the secretion of giant plasma membrane vesicles (or "blebs") (Scott, 1976; Baumgart et al. Because the spontaneous tension is a material parameter, tubulated vesicles behave, to a large extent, like liquid droplets with a variable surface area and with an effective interfacial tension that is provided by the spontaneous tension. This droplet-like behavior, which reflects the area reservoir that the nanotubes provide for the mother vesicle, leads to an increased robustness against mechanical perturbations as has been recently demonstrated by micropipette aspiration and cycles of osmotic deflation and inflation (Bhatia et al.
Order generic diamox on line
Definitive management Nonoperative management of pilon fractures is uncommon except for nondisplaced fractures (< 2 mm displacement with normal length and alignment) and in patients with prohibitive medical comorbidities or nonambulating patients medications not to be crushed buy diamox discount. Historically, definitive management should be delayed until soft-tissue swelling has subsided (positive wrinkle sign) and fracture blisters have sufficiently re-epithelialized; however, some studies have shown early definitive fixation may yield equivalent outcomes for select patients. Potential grafting or filling with bone substitutes of metaphyseal bone defects/loss. Most fractures are treated with plate and screw fixation; however, some type A and C1 (simple articular) fractures may be amenable to intramedullary fixation. The fibula is typically not fixed as this has been shown to have a higher complication rate with no clinical benefit. Both joint spanning frames and ring fixators (without spanning the joint) can be used. If a ring fixator is used, many times a temporary "foot plate" will be added initially and then removed 4 to 6 weeks post-op to allow early ankle range of motion. Medial to the tibialis anterior and lateral to the tribal crest proximal to the ankle joint. Lateral to the peroneus tertius/extensor digitorum communis and medial to the peroneal muscles in line with the fourth ray of the foot. Between the tibialis posterior tendon and flexor digitorum communis or between the flexor hallucis longus and the flexor digitorum communis. A, anteromedial; B, anterolateral; C, posterolateral fibula; D, posterolateral tibia, E, posteromedial tibia; F, medial tibia. A prospective study evaluating incision placement and wound healing for tibial plafond fractures. Note the external fixator was left in place due to significant anterior joint comminution in this case. Facilitates exposure of the fibula and the posterior malleolus for buttress plate application. Between the tibialis anterior and posterior tendons along the medial face of the tibia. Several strategies can be used to achieve this; however, two principles are frequently used for C-type fractures: a. This strategy can be employed when there is significant metaphyseal comminution with or without articular comminution. External fixators can be kept on for 6 weeks following fixation to neutralize deforming forces, help maintain length, and protect the articular reduction in the following circumstances: a. Definitive external fixation with a ring fixator, with or without limited internal fixation, might be considered for open pilon fractures, compartment syndrome, or patients with poor healing potential due to high risk of soft-tissue complications. Wound complication/dehiscence is relatively common and seen in up to 30% of patients. Often these can be treated with simple dressing changes but occasionally require return to the operating room for debridement. Post-traumatic arthritis is common, but patients may not require further surgery for several years. Conservative treatment options include bracing, nonsteroidal anti-inflammatory medication, acetaminophen, and cortisone injection. Reconstructive options for symptomatic post-traumatic arthritis that has failed conservative measures include fusion (most common), arthroplasty (rarely performed), and amputation in severe cases. Gentle range of motion exercises of the ankle can be started once the wound has healed. Staged management of most pilon fractures with meticulous soft-tissue management is vital in improving outcomes. The requirement of greater than 5 mm of displacement distinguishes pediatric pilon fractures from undisplaced triplane fractures. Be cognizant of the level of impaction that can occur in this patient population and the need for allograft augmentation. Consider leaving external fixation in place for 4 to 6 weeks postfixation if the patient has significant comminution and/or injury to the anterior aspect of the plafond that is causing subluxation of the talus anteriorly on the sagittal view. Summary Pilon fractures are complex fractures associated with significant high-energy soft tissue injuries. Optimal management requires thoughtful preoperative planning and careful management of soft tissues. Quality of life and prognostic factors after intra-articular tibial pilon fracture. Type C tibial pilon fractures: short- and long-term outcome following operative intervention. A staged protocol for soft tissue management in the treatment of complex pilon fractures. Schumaier Introduction Ankle fractures and fracture dislocations are the most frequent intra-articular fracture of the weight-bearing joints. Approximately 70% of ankle fractures are unimalleolar, 20% bimalleolar, and 10% trimalleolar (Video 43. Keywords: ankle, plafond, mortise, syndesmosis, malleolus, tibia, fibula, dislocation, fracture I. Mechanism of injury (typically rotational or abduction/adduction), ability to bear weight following injury, location(s) of pain, and prior injuries to ankle. Comorbid conditions are particularly relevant including diabetes mellitus, peripheral vascular disease, and preoperative ambulatory status. The soft-tissue envelope around the ankle can be tenuous, so definitive fixation may be delayed to allow soft-tissue recovery. Palpation: entire length of the tibia, fibula, and foot should be palpated for tenderness. Vascular examination: posterior tibial artery (posterior to the medial malleolus) and dorsalis pedis artery (lateral to the extensor hallucis longus tendon). Typically evaluate dorsal foot (superficial peroneal), plantar foot (tibial), and first web space (deep peroneal) sensation. Motor function is difficult to evaluate in a fracture situation, but assessing toe flexion and extension is usually possible. Fibula-slightly posterior to the tibia at the ankle; forms the lateral malleolus. Medial malleolus has an anterior and posterior colliculus with an intercollicular groove, where the deltoid ligament attaches. The dome is mostly covered with articular cartilage and is housed in the ankle mortise. Syndesmosis-composed of four ligaments between the distal tibia and fibula, which allow small amounts of motion. The superficial component connects the anterior colliculus of the edial malleolus m with the navicular, talus, and calcaneus. The deep component is mostly transverse and practically intra-articular, and connects the posterior colliculus of the medial malleolus to the talus. It is the strongest component and primary stabilizer, and can avulse the medial malleolus before tearing. Indications to obtain plain radiographs-history of frank dislocation, inability to bear weight following the injury, palpable tenderness of either malleolus. Mortise view-acquired by internally rotating the leg and foot 15 to 20 degrees until malleoli are equidistant from the image detector; it improves visualization of the joint space. External rotation and gravity stress views assess integrity of the deltoid ligament. Lateral collateral ligament-all the components attach to the lateral malleolus: i. Ankle fractures are typically described based on number of malleoli involved (unimalleolar, bimalleolar, trimalleolar).
Generic 250 mg diamox with visa
The two proteins catalyzed the synthesis of the two phospholipids medications vaginal dryness cheap 250 mg diamox overnight delivery, lysophosphatidic acid and phosphatidic acid. Unfortunately due to the low reaction yields and due to reaction network discontinuities, no morphological changes caused by the newly synthesized lipids could be observed. The amount of phospholipid products appeared to be restricted by the insufficient nutrient passage through the phospholipid membrane. In order to overcome such limitation in future, the vesicles must be a semi-open system as it is the case of living cells, that is, vesicle with chemically selective transport system. It stands somewhat between the simple fatty acid-based systems and the more sophisticated phospholipid vesicles with endovesicular protein expressions (Kurihara et al. This system was further advanced by investigating the recursive self-proliferation ability of the vesicles as protocell model system, which was, however, constructed from molecules that are not prebiotically plausible (Kurihara et al. All the efforts described here-and others described in the literature-are aimed at constructing minimal cells. If one is not too skeptical, then one may think that this type of experimental work and the general idea behind it may at some stage not only lead to the construction of synthetic minimal cells but it may also contribute to an understanding of the origin of the first cells. The first cells can be viewed as a particular type of minimal cells, but composed of chemically simpler molecules than contemporary cells are built from. The first cells must have contained only those molecules that were present on the early Earth due to nonbiological reactions. If the daughter vesicles have the same properties as the mother vesicle, the self-reproduction process can be repeated. Although many attempts have been made to realize the selfreproduction of vesicles using simple amphiphiles, the successes are still very limited, as described in Section 28. This fact suggests that the self-reproduction of vesicles is not straightforward from a membrane physics point of view (Svetina, 2012). The requirements to attain the self-reproduction of vesicles-independent of whether they are made from phospholipids, fatty acids, or other types of amphiphiles-are (i) vesicle deformation from a sphere to the limiting shape, (ii) breaking the neck of the limiting shape vesicle to produce daughter vesicles, and (iii) a recursive nature of the vesicles after division. The deformation of vesicles is well understood on the basis of the membrane elasticity energy model (curvature model) (Seifert, 1997), see also Chapter 5. The most successful model to describe vesicle shapes is the area-difference-elasticity model (Bozic et al. The area difference elastic energy originates from relative stretching of a monolayer. The bilayer has a preferred area difference given by A0 = (N + - N -) Alip, where Alip is the equilibrium area per membrane molecule and N+ and N- are the numbers of molecules in the outer and inner monolayer, respectively. In the case of "simple" vesicle systems (not containing complex endovesicular reactions), an understanding of the physics of the vesicles and their morphological changes may provide some useful insight (Sakuma and Imai, 2015), see also Chapter 5. This especially concerns aspects of (i) the self-reproduction of vesicles, and (ii) the exchange of nutrients from the outside to the inside of vesicles. These two topics are discussed in the following on the basis of a series of specifically 28. If the monolayer area difference A deviates from the preferred value A0, the monolayers should be stretched relative to one another, which gives the area difference elastic energy. In this model the vesicle shapes can be mapped by two geometrical parameters, the reduced volume expressed as: v= where Rs = V 4 Rs3 / 3 (28. In this phase diagram characteristic equilibrium shapes (pear, prolate, oblate, stomatocyte and sphere) and the limiting shape, are illustrated for each phase. The limiting shape line (L) and first-order discontinuous transitions, stomatocyte-oblate transition (Dsto/obl), oblate-prolate transition (Dobl/pro), and prolatepear transition (Dpro/pea) lines are indicated by solid lines. These transitions take place when the deformation trajectory a 0 (v) crosses the shape boundary lines. Thus incorporated membrane molecules should be delivered to the inner and outer leaflets to attain a0 1. Localization of the characteristic equilibrium shapes-sphere, pear, prolate, oblate, stomatocyte-and the limiting shape are illustrated for each phase. Solid lines indicate discontinuous transitions between the stomatocyte and the oblate shape (Dsto/obl), the oblate and the prolate shape (Dobl/pro), and the prolate and the pear shape (Dpro/pea). A unique technique to attain deformations to the limiting shape is to encapsulate colloidal particles (Natsume et al. By confining colloids densely, nonspherical vesicles deform to the limiting shape, see Chapters 8, 9, and 25. This deformation is caused by the maximization of the free volume of the confined colloids in a vesicle, that is, overlapping of the depletion zone next to the vesicle membrane (Asakura and Oosawa, 1954). The second issue concerns the mechanism of breaking the neck after the limiting shape has been reached (division process). It is well known that single-component vesicles can deform to the budded limiting shape in response to external stimuli. In such a case, vesicle division must involve a mechanism of neck destabilization, such as mechanical agitation (Zhu and Szostak, 2009; Adamala and Szostak, 2013). These experiments suggest that a coupling between Gaussian curvature and local lipid composition is important for such a division process. The free energy analysis for the limiting shape and the two-vesicle states shows that this coupling can destabilize the narrow neck in a limiting shape of binary vesicles, if the minor component lipids prefer to stay at regions with large positive Gaussian curvature (Chen et al. Thus the vesicle division is facilitated by mixing two types of amphiphiles having different Gaussian bending rigidities. In addition, after the neck has been broken, offspring vesicles should have the same properties (composition) as the mother vesicle so that the self-reproduction process can be repeated. Thus to attain vesicle self-reproduction, the vesicles should satisfy several physical requirements. After cooling back to the initial temperature, offspring vesicles recovered the original spherical shape of the mother vesicle. In the next cycle, the process was repeated in both offspring vesicles and then again, eventually yielding more than 10 generations of vesicles. The main paths for this transportation are (i) passive diffusion, (ii) pore formation, and (iii) fusion of cargo vesicles. Passive diffusion: the passive diffusion is governed by the permeability of the vesicle membrane. In the case of phospholipid vesicles, the permeability of small, uncharged molecules-such as water, oxygen, and carbon dioxide-is greater than the permeability of ionic compounds (Deamer and Dworkin, 2005; Monnard and Deamer, 2011) see also Chapter 20. This permeability is interpreted by the solubilitydiffusion model, where the permeability is simply governed by the hydrophobicity of the solute and the diffusion of the solute in the hydrophobic interior of the membrane. However, it was found that the permeability is strongly dependent on the chain length (Paula et al. For instance, shortening phospholipid chains from 18 to 14 carbons increases the permeability to ions by a thousand fold. To account for this observation, a transient pore model has been proposed (Paula and Deamer, 1999). The thinner membranes have increasing numbers of transient defects that open and close pores on nanosecond time scales. Using this property, vesicle systems with a temperature-controlled solute uptake (and release) can be designed. Compared with double-chain phospholipid vesicles, fatty acid vesicles have different membrane permeation properties because the neutral form of a single-chain fatty acid crosses a bilayer membrane with a half-time of milliseconds (flip-flop time), much faster than in the case of phospholipids (Mansy, 2009). The more dynamic fatty acid membranes were shown to be permeable remarkably to nucleoside mono- and diphosphates (Apel et al. Raising the temperature or adding low millimolar concentrations of Mg2+ further increases the permeability of fatty acid vesicles (Mansy and Szostak, 2008). In prebiotic times, where vesicular protocells may have existed, the passive diffusion might have been important for the uptake of critical nutrients. Pore formation: the formation of protein pores is a powerful means to transport chemical substances through vesicle membranes, like in the case of contemporary biomembranes. To form such pores, several pore-forming proteins and peptides can be utilized such as -hemolysin (Noireaux and Libchaber, 2004), porins (Graff et al. Pore formation can be achieved without the use of any proteins (or peptides) by coupling the spontaneous curvature of the lipids with lipid phase separation (Sakuma et al. The opening and closing of pores in a vesicle results from a competition between two forces: surface tension and line tension.
