

Discount 160mg fenofibrate amex
The churning action breaks the lipids into small droplets cholesterol from shrimp buy fenofibrate with mastercard, increasing the surface area for digestive enzymes. In the stomach, the lipid droplets are emulsified (kept apart) by dietary proteins. One of the most important contributions of the stomach to overall lipid digestion (and absorption) is that it empties chyme slowly into the small intestine, allowing adequate time for pancreatic enzymes to digest lipids. Phospholipase A2 is secreted as a proenzyme and, like many other pancreatic enzymes, is activated by trypsin. The final products of lipid digestion are monoglycerides, fatty acids, cholesterol, lysolecithin, and glycerol (from hydrolysis of ester bonds of triglycerides). With the exception of glycerol, each end product is hydrophobic and therefore is not soluble in water. Now the hydrophobic digestive products must be solubilized in micelles and transported to the apical membrane of the intestinal cells for absorption. Absorption of Lipids Most lipid digestion occurs in the small intestine, where conditions are more favorable than in the stomach. These bile salts, together with lysolecithin and products of lipid digestion, surround and emulsify dietary lipids. Emulsification produces small droplets of lipid dispersed in the aqueous solution of the intestinal lumen, creating a large surface area for the action of pancreatic enzymes. It hydrolyzes triglyceride molecules to one molecule of monoglyceride and two molecules of fatty acid. A potential problem in the action of pancreatic lipase is that it is inactivated by bile salts. Bile salts displace pancreatic lipase at the lipid-water interface of the emulsified lipid droplets. Colipase is secreted in pancreatic juices in an inactive form, procolipase, which is activated in the intestinal lumen by trypsin. Colipase then displaces bile salts at the lipid-water interface and binds to pancreatic lipase. With the inhibitory bile salts displaced, pancreatic lipase can proceed with its digestive functions. The products of lipid digestion (cholesterol, monoglycerides, lysolecithin, and free fatty acids) are solubilized in the intestinal lumen in mixed micelles, except glycerol, which is water soluble. As discussed earlier, the core of a micelle contains products of lipid digestion and the exterior is lined with bile salts, which are amphipathic. The hydrophilic portion of the bile salt molecules dissolves in the aqueous solution of the intestinal lumen, thus solubilizing the lipids in the micellar core. The micelles diffuse to the apical (brush-border) membrane of the intestinal epithelial cells. The micelles per se do not enter the cell, however, and the bile salts are left behind in the intestinal lumen to be absorbed downstream in the ileum. Because most ingested lipid is absorbed by the midjejunum, the "work" of the bile salts is completed long before they are returned to the liver via the enterohepatic circulation. Inside the intestinal epithelial cells, the products of lipid digestion are reesterified with free fatty acids on the smooth endoplasmic reticulum to form the original ingested lipids, triglycerides, cholesterol ester, and phospholipids. Inside the cells, the reesterified lipids are packaged with apoproteins in lipid-carrying particles called chylomicrons. Phospholipids cover 80% of the outside of the chylomicron surface, and the remaining 20% of the surface is covered with apoproteins. Apoproteins, which are synthesized by the intestinal epithelial cells, are essential for the absorption of chylomicrons. Failure to synthesize Apo B (or -lipoprotein) results in abetalipoproteinemia, a condition in which a person is unable to absorb chylomicrons and therefore is also unable to absorb dietary lipids. The secretory vesicles migrate to the basolateral membranes, and there is exocytosis of the chylomicrons. The chylomicrons are too large to enter vascular capillaries, but they can enter the lymphatic capillaries (lacteals) by moving between the endothelial cells that line the lacteals. The lymphatic circulation carries the chylomicrons to the thoracic duct, which empties into the bloodstream. Abnormalities of Lipid Digestion and Absorption the mechanisms for lipid digestion and absorption are more complex and involve more steps than those for carbohydrate and protein. Thus there are also more steps at which an abnormality of lipid digestion or absorption can occur. Each step in the normal process is essential: pancreatic enzyme secretion and function, bile acid secretion, emulsification, micelle formation, diffusion of lipids into intestinal epithelial cells, chylomicron formation, and transfer of chylomicrons into lymph. An abnormality at any one of the steps will interfere with lipid absorption and result in steatorrhea (fat excreted in feces). The gastric chyme, which is delivered to the duodenum, has a pH ranging from 2 at the pylorus to 4 at the duodenal bulb. The first reason is illustrated by Zollinger-Ellison syndrome, in which a tumor secretes large quantities of gastrin (Box 8. The elevated levels of gastrin stimulate excessive secretion of H+ by the gastric parietal cells, and this H+ is delivered to the duodenum, overwhelming the ability of pancreatic juices to neutralize it. Deficiency of bile salts interferes with the ability to form micelles, which are necessary for solubilization of the products of lipid digestion. Ileal resection (removal of the ileum) interrupts the enterohepatic circulation of bile salts, which then are excreted in feces rather than being returned to the liver. A 52-year-old man visits his physician complaining of abdominal pain, nausea, loss of appetite, frequent belching, and diarrhea. Because Zollinger-Ellison syndrome is suspected in this patient, his serum gastrin level is measured and found to be markedly elevated. While awaiting surgery, the man is treated with the drug omeprazole, which inhibits H+ secretion by gastric parietal cells. In Zollinger-Ellison syndrome, the tumor secretes large amounts of gastrin into the circulation. The target cell for gastrin is the gastric parietal cell, where it stimulates H+ secretion. The gastric G cells, the physiologic source of gastrin, are under negative feedback control. Thus normally, gastrin secretion and H+ secretion are inhibited when the gastric contents are acidified. In Zollinger-Ellison syndrome, however, this negative feedback control mechanism does not operate: Gastrin secretion by the tumor is not inhibited when the gastric contents are acidified. Therefore, gastrin secretion continues unabated, as does H+ secretion by the parietal cells. The presence of fat in the stool (steatorrhea) is abnormal because mechanisms in the small intestine normally ensure that dietary fat is completely absorbed. The duodenal contents remain at acidic pH rather than being neutralized, and the acidic pH inactivates pancreatic lipase. When pancreatic lipase is inactivated, it cannot digest dietary triglycerides to monoglycerides and fatty acids. Undigested triglycerides are not absorbed by intestinal epithelial cells, and thus they are excreted in the stool. The drug is expected to reduce H+ secretion and decrease the H+ load to the duodenum. Bacterial overgrowth reduces the effectiveness of bile salts by deconjugating them. In other words, bacterial actions remove glycine and taurine from bile salts, converting them to bile acids. Recall that at intestinal pH, bile acids are primarily in the non-ionized form (because their pKs are higher than intestinal pH); the non-ionized form is lipid soluble and readily absorbed by diffusion across the intestinal epithelial cells. For this reason, the bile acids are absorbed "too early" (before reaching the ileum), before micelle formation and lipid absorption are completed. Similarly, decreased pH in the intestinal lumen promotes "early" absorption of bile acids by converting them to their non-ionized form.
Purchase cheap fenofibrate line
For example cholesterol free shrimp fenofibrate 160 mg online, if the total blood volume is 5 L, 4 L is in the unstressed volume, producing no pressure, and 1 L is in the stressed volume, producing a pressure of approximately 7 mm Hg (on the graph, read mean systemic pressure as 7 mm Hg at a blood volume of 5 L). If blood volume increases, the amount of blood in the unstressed volume will be unaffected (if it is already full), but the amount of blood in the stressed volume will increase. When stressed volume increases, mean systemic pressure increases and the vascular function curve and its intersection point with the X-axis shift to the right. If blood volume decreases, then stressed volume decreases, mean systemic pressure decreases, and the vascular function curve and its intersection point with the X-axis shift to the left. Redistribution of blood between the unstressed volume and the stressed volume also produces changes in mean systemic pressure. Although total blood volume is unchanged, the shift of blood increases the mean systemic pressure and shifts the vascular function curve to the right. Hence, the unstressed volume will increase, the stressed volume and mean systemic pressure will decrease, and the vascular function curve shifts to the left. In summary, increased blood volume and decreased compliance of the veins produce an increase in mean systemic pressure and shift the vascular function curve to the right. Decreased blood volume and increased compliance of the veins produce a decrease in mean systemic pressure and shift the vascular function curve to the left. Total blood volume is the sum of unstressed volume (in the veins) and stressed volume (in the arteries). If mean systemic pressure is fixed or constant, the slope of the vascular function curve can be changed by rotating it. A clockwise rotation means that, for a given right atrial pressure, venous return is increased. A counterclockwise rotation means that, for a given right atrial pressure, venous return is decreased. The point at which the two curves intersect is the unique operating or equilibrium point of the system in the steady state. In the steady state, cardiac output and venous return are, by definition, equal at the point of intersection. Why then do the cardiac and vascular function curves go in opposite directions and why do they have opposite relationships with right atrial pressure The cardiac function curve is determined as follows: As right atrial pressure and end-diastolic volume are increased, there is increased ventricular fiber length, which leads to increased stroke volume and cardiac output. The higher the right atrial pressure, the higher the cardiac output-this is the Frank-Starling relationship for the heart. The vascular function curve is determined as follows: As right atrial pressure is decreased, venous return increases because of the greater pressure gradient driving blood flow back to the heart. We have established that cardiac and vascular function curves have opposite relationships with right atrial pressure. At this one value of right atrial pressure, cardiac output equals venous return and, by definition, is the steady state operating point of the system. That one value of right atrial pressure satisfies both cardiac output and venous return relationships. Combining these curves provides a useful tool for predicting the changes in cardiac output that will occur when various cardiovascular parameters are altered. Cardiac output can be altered by changes in the cardiac function curve, by changes in the vascular function curve, or by simultaneous changes in both curves. The basic premise of this approach is that, after such a change, the system will move to a new steady state. In the new steady state, the operating point at which the cardiac and the vascular function curves intersect will have changed. This new operating point tells what the new cardiac output and the new venous return are in the new steady state. Recall that positive inotropic agents cause an increase in contractility for a given end-diastolic volume (or right atrial pressure), and negative inotropic agents produce a decrease in contractility. Positive inotropic agents produce an increase in contractility, an increase in stroke volume, and an increase in cardiac output for any level of right atrial pressure. Thus the cardiac function curve shifts upward, but the vascular function curve is unaffected. The point of intersection (the steady state point) of the two curves now has shifted upward and to the left. In the new steady state, cardiac output is increased and right atrial pressure is decreased. The decrease in right atrial pressure reflects the fact that more blood is ejected from the heart on each beat as a result of the increased contractility and increased stroke volume. The effect is just the opposite of a positive inotropic agent: There is a decrease in contractility and a decrease in cardiac output for any level of right atrial pressure. The cardiac function curve shifts downward, and the vascular function curve is unchanged. In the new steady state, cardiac output is decreased and right atrial pressure is increased. Right atrial pressure is increased because less blood is ejected from the heart on each beat, due to decreased contractility and decreased stroke volume. The circle intersecting the dashed line shows the new steady state operating point. Increases in blood volume increase the amount of blood in the stressed volume and therefore increase the mean systemic pressure. Mean systemic pressure is the point on the vascular function curve where venous return is zero. Increases in blood volume shift this intersection point to the right and therefore shift the curve to the right in a parallel manner. The decrease in blood volume decreases the amount of blood in the stressed volume and mean systemic pressure, which shifts the vascular function curve to the left in a parallel manner. In the new steady state, cardiac output is decreased and right atrial pressure is decreased. Changes in venous compliance produce effects similar to those produced by changes in blood volume. Decreases in venous compliance cause a shift of blood out of the unstressed volume and into the stressed volume and produce changes similar to those caused by increases in blood volume, a parallel shift to the right. Likewise, increases in venous compliance cause a shift of blood into the unstressed volume and out of the stressed volume and produce changes similar to those caused by decreased blood volume, a parallel shift to the left. This increase in arterial pressure produces an increase in afterload on the heart, which decreases cardiac output. The cardiac function curve shifts downward as a result of the increased afterload. This rotation means that less blood returns to the heart for a given right atrial pressure- venous return is decreased. The curves intersect at a new steady state point at which both cardiac output and venous return are decreased. Such changes alter the extent to which blood is "held" on the arterial side of the circulation. The solid lines show the normal relationships, and the dashed lines show the changes. The circle intersecting the dashed lines shows the new steady state operating point. Depending on the relative magnitude of the effects on the cardiac and vascular function curves, right atrial pressure can be slightly increased, slightly decreased, or unchanged. The curves intersect at a new steady state point at which both cardiac output and venous return are increased. Depending on the relative magnitude of the effects, right atrial pressure can be slightly increased, slightly decreased, or unchanged. The mechanisms that help to maintain Pa at a constant value are discussed in this section. These mechanisms closely monitor Pa and compare it with the set-point value of approximately 100 mm Hg. The baroreceptor reflex attempts to restore Pa to its set-point value in a matter of seconds. The second system is hormonally mediated and includes the reninangiotensin-aldosterone system, which regulates Pa more slowly, primarily by its effect on blood volume. Blood flow to the tissues is driven by the difference in pressure between the arterial and venous sides of the circulation.
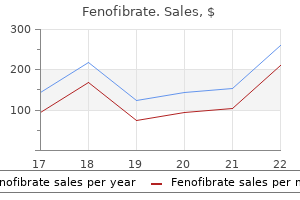
Purchase fenofibrate 160mg otc
The cells were then dissociated cholesterol/hdl ratio guidelines buy cheap fenofibrate line, labeled with tritiated thymidine, and injected into a host cortex at a different stage of development. Thus, the early-born neurons could respond to environmental cues in the host environment and migrate to a new destination. This effect was only seen, however, when the cells were harvested early in the cell cycle, prior to the final mitotic division. This finding suggested there were also intrinsic temporal cues present to influence fate options of cortical neurons. Thus, by the time a cell is post-mitotic, cell fate is established and cannot be altered, even when placed in a new host environment. A third set of experiments confirmed that the fate potential of the cortical neurons becomes gradually restricted during normal development. Together, the transplantation experiments revealed early-mid-stage progenitors are multipotent early in the cell cycle and can adopt new cortical fates when placed in an older host environment. As cells reach mid-late stages of development they become restricted in cell fate options. Thus, the progenitors arising from mid-late stages of cortical development are unable to adopt the fates of younger progenitors and remain committed to the fate corresponding to their time of migration. A number of subsequent studies have identified transcription factors that are specific to neurons located in different cortical layers. Thus, early-stage cortical progenitors are able to alter their fate and migrate to a new layer. However, this effect was only seen if the cells were harvested early in the cell cycle, prior to the final mitotic division. It has been suggested that some combinations of transcription factors are mutually repressive to prevent cells from adopting the fate of cells in adjacent layers. Ongoing studies seek to identify how transcription factors determine cortical layer cell fate. In at least some cases, the identified transcription factors regulate differentiation of neurons once a layer-specific fate has been established. Recent evidence has also indicated that temporal identity factors homologous to those found in Drosophila play a role in cerebral cortical fate potential. For example, Ikaros, the mammalian ortholog of hunchback, is expressed in early-stage cortical progenitors. In mice, Ikaros is detected in early-stage progenitors of the ventricular zone, but is decreased at later stages. When Ikaros was overexpressed in mice, the number of progenitor neurons was increased. If Ikaros was misexpressed in later-born progenitors, early-born fates could not be generated, consistent with the idea that Ikaros encodes a temporal factor utilized only by early-generated progenitor cells, similar to the function of hunchback in Drosophila neuroblasts. Ikaros appears to provide the early-generated neurons with the ability to adopt deep-layer cortical fates. The expression of other transcription factors is then needed for the cells to differentiate into mature cortical neurons with the characteristics typical of cells in that layer. Epigenetic factors influence determination and differentiation in vertebrate neurons In recent decades studies have also begun to focus on how epigenetic factors influence cell fate options in the developing nervous system. However, if Ikaros was expressed in later-born progenitors, cell fate was not altered (not shown). These results support the hypothesis that the temporal identify factor Ikaros, like Hunchback, only influences the fate of early-generated progenitor cells. For example, when chromatin is in a lightly packed (euchromatin) state, the corresponding promoter region of a target gene becomes accessible to transcription factors. Brg1 appears to be particularly important during neural proliferation, whereas Brm is required for cell fate determination of progenitors and differentiation of post-mitotic neurons. Brg1 and other subunits are needed to maintain Notch signaling in order to repress proneural genes and keep cells in a proliferative state. In contrast, Brm and other subunits activate the transcription of neuron-specific genes such as Neurogenin and NeuroD. The changes in subunit composition correlate with the transition to each stage of neuronal development. The changes in the expression of these subunits is correlated with the transition from proliferating progenitors to committed cortical and granule cell fates. In the cerebral cortex, Pax6 activates target genes such as Tbr2 (T-box brain 2), which is detected in nonproliferating progenitors (for example, the basal progenitor cells). Pax 6 then activates Cux1, which is detected in post-mitotic neurons, and Tle1 (transducin-like enhancer protein-1), which is needed for the survival of post-mitotic neurons. Thus, Pax6 cannot readily bind to the target genes and initiate their expression in proliferating cells. These examples indicate one of the ways epigenetic regulation of transcription factor binding sites can influence whether genes necessary for determination and subsequent differentiation are expressed. Thus, epigenetic modifications provide another means by which the limited number of available transcription factors can exert a wide range of effects in the developing nervous system. As discussed in Chapter 5, the fate options available to neural crest cells are probably the most varied in the nervous system, and each neural crest cell population relies on specific signals for determination and differentiation. Most neural crest cells appear to be particularly influenced by extrinsic signals encountered as they migrate from the neural folds toward their final destinations. Environmental cues influence the fate of parasympathetic and sympathetic neurons Neural crest cells from the caudal hindbrain through the sacral region are divided into vagal and trunk populations. Among the derivatives of the vagal and trunk neural crest cells are neurons in the parasympathetic and sympathetic divisions of the autonomic nervous system. In contrast, sympathetic neurons that innervate smooth muscle cells and utilize the neurotransmitter norepinephrine (also called noradrenaline) are derived from the trunk neural crest. The vagal and trunk populations of neural crest cells have been utilized extensively to evaluate whether neural crest cell fate is predetermined or regulated by cues from the extracellular environment. The influence of the environment on the fate options of parasympathetic and sympathetic neurons from the vagal and trunk regions of the neural tube was first described in the now-classic studies of Nicole LeDouarin and colleagues in the 1970s. These studies relied on transplantation techniques pioneered by LeDourain in which the neural crest cells of quail were transplanted to a chick embryo at a similar stage of development. In such cases, the quail cells integrate into the chick host and differentiate as if they were chick cells. Because neural crest development occurs in a rostral-to-caudal progression, cells from the vagal region develop prior to those of the trunk region. Histological differences between quail and chick cells allowed researchers to trace the fate of donor cells in the resulting chimeras. The size difference between the quail donors and chick hosts reflects differences in the developmental stage of the embryos. Development in the vagal region precedes trunk development, so to ensure that the transplanted donor cells are at the same developmental stage as their neighboring host cells, cells from the vagal region of a younger donor were transplanted into the trunk region of an older host (A), whereas cells from the trunk region of an older donor were transplanted into the vagal region of a younger host (B). Similarly, when trunk neural crest cells were transplanted to vagal regions, the trunk cells took the expected migratory route for vagal crest cells and became parasympathetic neurons. Thus, the fate of parasympathetic and sympathetic neurons was not predetermined, but appeared to depend on cues encountered along the migratory route of the neural crest cells. Experiments have also indicated that the differences in fate options do not result from the selective survival of neural crest cells in different regions, but are largely caused by environmental cues produced in different tissues at each axial level. Thus, it appears that neural crest cells arise from a multipotent precursor population that can give rise to a number of different cell types. More recent studies have supported the finding that environmental cues can alter neural crest fate by regulating the expression of specific transcription factors, at least during defined stages of development. These Ngns then activate the expression of the neuronal differentiation marker, NeuroD. Cells expressing Ngns also increase their expression of a Notch ligand called Delta-like ligand 1 (Dll1). Dll1 binds to Notch receptors expressed on surrounding cells, thus preventing those cells from developing as neurons. As a result, only those cells expressing Ngns will be able to adopt a neural fate. The majority of sympathetic neurons go on to innervate tissues such as skin (A) All sympathetic neurons are initially adrenergic, producing the neurotransmitter norepinephrine (also called noradrenaline).
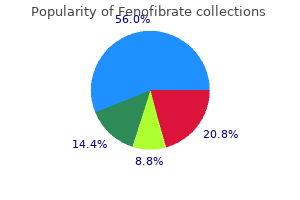
Fenofibrate 160mg mastercard
In the vagina cholesterol levels very high purchase 160 mg fenofibrate mastercard, estrogen stimulates proliferation of epithelial cells; progesterone stimulates differentiation but inhibits proliferation of epithelial cells. Based on a "typical" 28-day cycle, the follicular phase of the menstrual cycle is the 14-day period preceding ovulation. This phase, which is also called the proliferative phase, is dominated by estrogen. When spread on a glass slide, cervical mucus from the follicular phase produces a pattern known as "ferning. The luteal phase of a 28-day menstrual cycle is the 14-day period following ovulation. The uterine glands become more tortuous, accumulate glycogen in vacuoles, and increase their mucus secretions. Progesterone secretion decreases the quantity of cervical mucus, which then becomes thick and nonelastic and does not "fern" on a slide. During this period, a primordial follicle develops into a graafian follicle and neighboring follicles become atretic (degenerate or regress). After the neighboring follicles degenerate, the remaining follicle is called the dominant follicle. The follicular phase is dominated by 17-estradiol, whose levels steadily increase. For example, in a 35-day cycle, ovulation occurs on day 21, or 14 days before menses; in a 24-day cycle, ovulation occurs on day 10. At ovulation, cervical mucus increases in quantity and becomes watery and more penetrable by sperm. Estradiol levels decrease just after ovulation, but they will increase again during the luteal phase. During the luteal phase, the corpus luteum develops and begins synthesizing estradiol and progesterone. The high levels of progesterone during this phase stimulate secretory activity of the endometrium and increase its vascularity. Thus in the follicular phase, estradiol causes the endometrial lining to proliferate; in the luteal phase, progesterone is preparing the endometrium to receive a fertilized ovum. Basal body temperature increases during the luteal phase because progesterone increases the hypothalamic temperature set point. The cervical mucus becomes less abundant and thicker, and it is now "too late" for sperm to fertilize the ovum. Late in the luteal phase, if fertilization has not occurred, the corpus luteum regresses. With this regression, the luteal source of estradiol and progesterone is lost, and blood levels of the hormones decrease abruptly. The breasts, or mammary glands, are composed of lobular ducts lined by a milk-secreting epithelium. At puberty, with the onset of estrogen secretion, the lobular ducts grow and the area around the nipple, the areola, enlarges. Estrogen also increases the amount of adipose tissue, giving the breasts their characteristic female shape. Progesterone collaborates with estrogen by stimulating secretory activity in the mammary ducts. Pregnancy the highest levels of estrogen and progesterone occur during pregnancy, synthesized in early pregnancy by the corpus luteum and in mid-to-late pregnancy by the placenta. Estrogen stimulates growth of the myometrium, growth of the ductal system of the breasts, prolactin secretion, and enlargement of the external genitalia. Progesterone maintains the endometrial lining of the uterus and increases the uterine threshold to contractile stimuli, thus preserving the pregnancy until the fetus is ready to be delivered. Other Actions of Estrogen and Progesterone In addition to those actions previously discussed, estrogen contributes to the pubertal growth spurt, closure of the epiphyses at the end of the growth spurt, and the deposition of subcutaneous fat. Progesterone has a mild thermogenic action, which increases basal body temperature during the luteal phase of the menstrual cycle. This increase in basal body temperature during the luteal phase is the basis for the "rhythm" method of contraception, in which the increase in temperature can be used retrospectively to determine the time of ovulation. Events of the Menstrual Cycle the menstrual cycle recurs approximately every 28 days over the reproductive period of the female: from puberty until menopause. The events of the cycle include development of an ovarian follicle and its oocyte, ovulation, preparation of the reproductive tract to receive the fertilized ovum, and shedding of the endometrial lining if fertilization does not occur. The variability in cycle length is attributable to variability in the duration of the follicular phase; the luteal phase is constant. Regression of the corpus luteum and the abrupt loss of estradiol and progesterone cause the endometrial lining and blood to be sloughed (menses or menstrual bleeding). During this time, primordial follicles for the next cycle are being recruited and are beginning to develop. Pregnancy If the ovum is fertilized by a sperm, the fertilized ovum begins to divide and will become the fetus. The period of development of the fetus is called pregnancy or gestation, which, in humans, lasts approximately 40 weeks. Their functions include maintenance of the endometrium, development of the breasts for lactation after delivery, and suppression of the development of new ovarian follicles. In early pregnancy (the first trimester), the source of steroid hormones is the corpus luteum; in mid-to-late pregnancy (the second and third trimesters), the source is the placenta. The blastocyst floats freely in the uterine cavity for 1 day and then implants in the endometrium 5 days after ovulation. The receptivity of the endometrium to the fertilized ovum is critically dependent on a low estrogen/progesterone ratio and corresponds to the period of highest progesterone output by the corpus luteum. At the time of implantation, the blastocyst consists of an inner mass of cells, which will become the fetus, and an outer rim of cells called the trophoblast. The trophoblast invades the endometrium and forms an attachment to the maternal membranes. At the point of implantation, under stimulation by progesterone, the endometrium differentiates into a specialized layer of decidual cells. Trophoblastic cells proliferate and form the syncytiotrophoblast, whose function is to allow the blastocyst to penetrate deep into the endometrium. The timetable is based on the number of days after ovulation and includes the following steps: 1. Fertilization of the ovum takes place within 24 hours of ovulation, in a distal portion of the oviduct called the ampulla. Once a sperm penetrates the ovum, the second polar body is extruded and the fertilized ovum begins to divide. Four days after ovulation the fertilized ovum, the blastocyst, with approximately 100 cells, arrives in the uterine cavity. Days After Ovulation 0 day 1 day 4 days 5 days 6 days 8 days 10 days the duration of pregnancy is, by convention, counted from the date of the last menstrual period. Pregnancy lasts approximately 40 weeks from the onset of the last menstrual period, or 38 weeks from the date of the last ovulation. Pregnancy is divided into three trimesters, each of which corresponds to approximately 13 weeks. Parturition Pr og E io str l Parturition, the delivery of the fetus, occurs approximately 40 weeks after the onset of the last menstrual period. The mechanism of parturition is unclear, although roles for estrogen, progesterone, cortisol, oxytocin, prostaglandins, relaxin, and catecholamines have been proposed. Once the fetus reaches a critical size, distention of the uterus increases its contractility. Uncoordinated contractions, known as Braxton Hicks contractions, begin approximately 1 month before parturition. Near term, the fetal hypothalamic-pituitary-adrenal axis is activated and the fetal adrenal cortex produces significant amounts of cortisol. Cortisol increases the estrogen/progesterone ratio, which increases the sensitivity of the uterus to contractile stimuli.
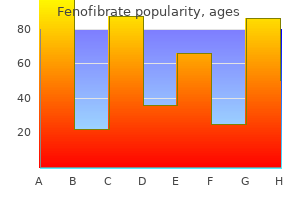
Order fenofibrate online pills
Different proteins associated with endosomes at early and late stages of transport have been noted cholesterol levels by age generic 160mg fenofibrate visa. Once the endosome complex reaches the cell body, intracellular signaling cascades, such as those described below, are initiated to promote cell survival. Transport requirements for neurotrophin signaling remain an ongoing focus of investigation. The ligand remains inside the endosome, while the receptor kinase domains extend outward from the vesicle surface. Because the ligand is still bound to the receptor, the receptor remains active during transport and recruits additional signaling molecules to the endosome surface. Once the endosome complex reaches the cell body, it is able to initiate signal transduction cascades associated with cell survival. These phosphorylated sites then recruit other proteins that initiate intracellular signaling cascades. Among the proteins that bind to phosphorylated sites on the Trk receptor are adapter proteins such as Shc (C-terminal dn 8. These proteins, in turn, activate molecular pathways leading to various transcriptional events that regulate functions as varied as neurite outgrowth and synaptic plasticity, cell differentiation, and neuronal survival. The precise intracellular effects of the neurotrophins depend in part on the cellular context and stage of development. These pathways often interact through additional proteins, however, and overlapping functions are found in several neuronal contexts. Ras pathway activation is initiated with the recruitment of the adaptor protein Shc to the phosphorylated tyrosine residues on the receptor. Upon neurotrophin binding, phosphorylation of kinase residues outside of the kinase domains recruits adaptor proteins to the receptor, thereby initiating various signal transduction cascades. Phosphorylated Erk enters the nucleus, where it regulates various genes associated with neuronal differentiation. Activation of either ultimately influences the expression of genes associated with neuronal survival. In addition, Akt can inhibit cell death pathways, as discussed later in the chapter. Although the function of these receptors remains largely unclear, studies suggest truncated TrkB and TrkC may sequester their corresponding neurotrophins to increase the local concentration of those growth factors, then somehow direct the ligands to full-length receptors on the cells that require them. Alternatively, the truncated receptors may take up and sequester growth factors to prevent activation of a full-length Trk receptor. However, neurotrophins are not the only family of molecules known to influence developing neurons. However, not all of these proteins meet the criteria for targetderived growth factors as defined by the original neurotrophic hypothesis. In 1979, Silvio Varon and colleagues isolated a survival- and outgrowth-promoting factor from the ciliary body and choroid of embryonic chick eye. This hypothesis was supported by studies in which the axons of facial motor neurons were severed (axotomized). Another difficulty is diagnosing motor neuron disease early enough to deliver growth factors before neuronal damage is too advanced to be reversed. Investigators also continue to identify the target-derived signals required for embryonic motor neurons. Early studies by Hamburger and others demonstrated that motor neuron survival was influenced by the size of available limb bud tissue, yet the growth factor that regulates motor neuron survival during development is still unknown. Several identified growth factors meet some of the necessary criteria to be considered a target-derived factor for motor neurons. In addition to growth factors present in skeletal muscle, many growth factors are expressed in the Schwann cells surrounding the peripheral axons of motor neurons. These are believed to be an additional source of survival-promoting factors for innervating motor neurons. The number of candidate growth factors identified for motor neurons has been referred to as "an embarrassment of riches. Thus, over the course of evolution, cells have developed and maintained mechanisms to induce cell death. That cells would expend energy to do this suggests there must be some evolutionary advantage to what may at first seem to be a wasteful process. During necrotic cell death, the dying cell begins to swell and develop cytoplasmic vacuoles. The nucleus and organelles then begin to break down, and as the cells continue to expand, the cell membrane eventually ruptures. Because macrophages become active when the cellular contents are released, an inflammatory response occurs in response to necrotic cell death. During apoptotic cell death, the cell cytoplasm shrinks, causing the cells to appear smaller. When examined under the microscope, an apoptotic cell reveals a pyknotic nucleus with highly condensed nuclear chromatin. The nucleus and organelles form distinct, membrane-bound fragments termed apoptotic bodies that are a hallmark of apoptosis. These apoptotic bodies are then phagocytized by macrophages to remove cellular debris. In some cases, glial cells adjacent to the apoptotic cells will function as macrophages to remove the apoptotic bodies. There is no inflammatory response to apoptosis, because phagocytosis occurs before the dying cells can release their contents. The characteristic changes that occur during apoptosis have been used to develop laboratory techniques that readily identify apoptotic cells. The breaks occur between nucleosomes creating fragments that are approximately 180 base pairs (bp) or multiples of 180 bp in length (that is, 360 bp, 540 bp, 720 bp, and so on). When a normal cell dies, the stages of cell death differ depending on whether the cell undergoes necrosis or apoptosis. At the onset of necrotic death, the cell begins to swell, and vacuoles form in the cytoplasm and organelles. The cell continues to enlarge as the nuclei and organelles disintegrate, until the cell membrane ultimately ruptures. As the cell initiates apoptotic death, its cytoplasm and nuclear chromatin begin to condense. At first, the organelles and cell membrane remain intact, even as the cell body and nucleus begin to shrink. However, the cell surface soon becomes convoluted, and the nucleus and organelles break off into discrete, membranebound fragments called apoptotic bodies. These apoptotic bodies are degraded and ultimately phagocytized by nearby macrophages or glial cells. The size of the bands produced is determined by comparing them to marker standards that are run at the same time. Markers standards (1) are run on the gel to estimate the size of the bands detected in the cell lysates. Cells that do not undergo apoptotic cell death do not reveal a ladderlike pattern (2). Scientists cultured sympathetic neurons harvested at the time of normal target innervation to test whether protein synthesis was a necessary step in the induction of cell death. The comparatively small number of cells found in specific locations helps scientists track what becomes of individual cells, including neurons. Several genes discovered in the 1990s were found to prevent cell death when mutated.
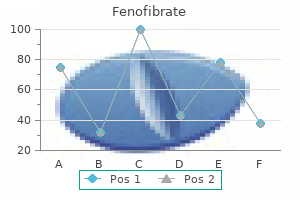
Purchase cheapest fenofibrate and fenofibrate
Ca2+ then binds to troponin C to allow actin-myosin interaction and cross-bridge formation cholesterol levels hdl ldl purchase fenofibrate 160 mg with mastercard. Inotropism or contractility is the ability of the myocardial cell to develop tension at a given cell length: Intracellular [Ca2+] determines the degree of inotropism, with positive inotropic agents increasing intracellular [Ca2+] and contractility. Myocardial cells and the myocardium exhibit a length-tension relationship based on the degree of overlap of contractile elements. The Frank-Starling law of the heart describes this relationship between cardiac output and end-diastolic volume. Therefore cardiac output is determined by venous return, and in the steady state, cardiac output and venous return are equal. The baroreceptor reflex is a fast, neural mechanism that detects changes in Pa and orchestrates changes in sympathetic and parasympathetic outflow to the heart and blood vessels to restore Pa back to normal. The exchange of fluid across capillary walls is determined by the balance of Starling forces. The net Starling pressure determines whether there will be filtration out of the capillary or absorption into the capillary. If filtration of fluid exceeds the ability of the lymphatics to return it to the circulation, then edema occurs. The blood flow to the organ systems is a variable percentage of the cardiac output. Blood flow is determined by arteriolar resistance, which can be altered by vasodilator metabolites or by sympathetic innervation. How much does total resistance change if a fourth resistor with a value of 10 is added in parallel If blood flow through the two vessels is the same, in which vessel is velocity of blood flow higher For each item in the following list, give its correct location in the cardiovascular system. The location may be anatomic, a graph or portion of a graph, an equation, or a concept. Dicrotic notch 1 receptors Lmax Radius to the fourth power Phospholamban Negative dromotropic effect Pulse pressure Normal automaticity Ejection fraction 25 During which portions(s) of the cardiac cycle is the mitral valve closed: atrial systole, rapid ventricular ejection, isovolumetric ventricular relaxation, diastasis Fresh air is brought into the lungs during the inspiratory phase of the breathing cycle, oxygen and carbon dioxide are exchanged between inspired air and pulmonary capillary blood, and the air is then expired. The structures of the respiratory system are subdivided into a conducting zone (or conducting airways), which brings air into and out of the lungs, and a respiratory zone lined with alveoli, where gas exchange occurs. Conducting Zone the conducting zone includes the nose, nasopharynx, larynx, trachea, bronchi, bronchioles, and terminal bronchioles. These structures function to bring air into and out of the respiratory zone for gas exchange and to warm, humidify, and filter the air before it reaches the critical gas exchange region. The trachea divides into the right and left mainstem bronchi (the first generation), which divide into two smaller bronchi, which divide again. Ultimately, there are 23 such divisions into increasingly smaller airways, culminating in the airways of the 23rd generation. The conducting airways are lined with mucus-secreting and ciliated cells that function to remove inhaled particles. Although large particles usually are filtered out in the nose, small particles may enter the airways, where they are captured by mucus, which is then swept upward by the rhythmic beating of the cilia. This smooth muscle has both sympathetic and parasympathetic innervations, which have opposite effects on airway diameter: (1) Sympathetic adrenergic neurons activate 2 receptors on bronchial smooth muscle, which leads to relaxation and dilation of the airways. Changes in diameter of the conducting airways result in changes in their resistance, which produce changes in air flow. Thus the effects of the autonomic nervous system on airway diameter have predictable effects on airway resistance and air flow. Respiratory Zone the respiratory zone includes the structures that are lined with alveoli and therefore participate in gas exchange: the respiratory bronchioles, alveolar ducts, and alveolar sacs. Like the conducting airways, they have cilia and smooth muscle, but they also are considered part of the gas exchange region because alveoli occasionally bud off their walls. The alveolar ducts are completely lined with alveoli, but they contain no cilia and little smooth muscle. The alveoli are pouchlike evaginations of the walls of the respiratory bronchioles, the alveolar ducts, and the alveolar sacs. Macrophages fill with debris and migrate to the bronchioles, where the beating cilia carry debris to the upper airways and the pharynx, where it can be swallowed or expectorated. Pulmonary Blood Flow Pulmonary blood flow is the cardiac output of the right heart. The pulmonary arteries branch into increasingly smaller arteries and travel with the bronchi toward the respiratory zones. The smallest arteries divide into arterioles and then into the pulmonary capillaries, which form dense networks around the alveoli. Because of gravitational effects, pulmonary blood flow is not distributed evenly in the lungs. When a person is standing, blood flow is lowest at the apex (top) of the lungs and highest at the base (bottom) of the lungs. The physiologic significance of regional variations in blood flow is discussed later in the chapter. As in other organs, regulation of pulmonary blood flow is accomplished by altering the resistance of the pulmonary arterioles. Changes in pulmonary arteriolar resistance are controlled by local factors, mainly O2. Bronchial circulation is the blood supply to the conducting airways (which do not participate in gas exchange) and is a very small fraction of the total pulmonary blood flow. Lung Capacities In addition to these lung volumes, there are several lung capacities; each lung capacity includes two or more lung volumes. Its value increases with body size, male gender, and physical conditioning and decreases with age. In the helium dilution method, the subject breathes a known amount of helium, which has been added to the spirometer. Because helium is insoluble in blood, after a few breaths the helium concentration in the lungs becomes equal to that in the spirometer, which can be measured. The amount of helium that was added to the spirometer and its concentration in the lungs are used to "back-calculate" the lung volume. Therefore if volume increases, pressure must decrease, and if volume decreases, pressure must increase. Typically, the subject is sitting and breathes into and out of the spirometer, displacing a bell. Normal tidal volume is approximately 500 mL and includes the volume of air that fills the alveoli plus the volume of air that fills the airways. Next, the subject is asked to take a maximal inspiration, followed by a maximal expiration. The additional volume that can be inspired above tidal volume is called the inspiratory reserve volume, which is approximately 3000 mL. The additional volume that can be expired below tidal volume is called the expiratory reserve volume, which is approximately 1200 mL. Simultaneously, the volume in the box decreases, and the pressure in the box increases. Dead Space Dead space is the volume of the airways and lungs that does not participate in gas exchange. Dead space is a general term that refers to both the anatomic dead space of the conducting airways and a functional, or physiologic, dead space. The next air to enter the alveoli is fresh air from the inspired tidal volume (350 mL), which will undergo gas exchange. The rest of the tidal volume (150 mL) does not make it to the alveoli but remains in the conducting airways; this air will not participate in gas exchange and will be the first air expired. Thus for example, when a tidal volume of 500 mL is inspired, the entire volume does not reach the alveoli for gas exchange; 150 mL fills the conducting airways (the anatomic dead space, where no gas exchange occurs), and 350 mL fills the alveoli. With the inspiration of the next tidal volume, this alveolar air is first to enter the alveoli, although it will not undergo further gas the concept of physiologic dead space is more abstract than the concept of anatomic dead space. By definition, the physiologic dead space is the total volume of the lungs that does not participate in gas exchange.
Diseases
- Pentosuria
- Wohlwill Andrade syndrome
- Anaplasmosis
- Proconvertin deficiency, congenital
- Tollner Horst Manzke syndrome
- Cocaine antenatal infection
- Coloboma of iris
Fenofibrate 160mg low price
Using the acid-base map cholesterol lowering foods eggs purchase 160mg fenofibrate visa, it can be concluded that the patient has chronic respiratory acidosis. The compensation is exactly as predicted by the rules of thumb for chronic respiratory acidosis. It can be concluded that the patient has simple chronic respiratory acidosis with the expected level of renal compensation. The difference between acute and chronic respiratory alkalosis lies in renal compensation. The mechanisms that maintain a constant pH include buffering, respiratory compensation, and renal compensation. When there is a gain of fixed H+, metabolic acidosis occurs; when there is a loss of fixed H+, metabolic alkalosis occurs. Which of the following might cause this pattern: diarrhea, vomiting, obstructive pulmonary disease, hysterical hyperventilation, salicylate overdose, chronic renal failure One has acute respiratory acidosis and the other has chronic respiratory acidosis. To serve these functions, there are four major activities of the gastrointestinal tract. The rate at which food is propelled through the gastrointestinal tract is regulated to optimize the time for digestion and absorption. Other structures of the gastrointestinal tract are the salivary glands, pancreas, liver, and gallbladder, all of which serve secretory functions. The layers of the gastrointestinal wall are as follows, starting from the lumen and moving toward the blood: A mucosal layer consists of a layer of epithelial cells, a lamina propria, and a muscularis mucosae. The epithelial cells are specialized to carry out absorptive and secretory functions. The lamina propria consists primarily of connective tissue, but it also includes blood and lymph vessels. The muscularis mucosae consists of smooth muscle cells; contraction of the muscularis mucosae changes the shape and surface area of the epithelial cell layer. Beneath the mucosal layer is a submucosal layer, which consists of collagen, elastin, glands, and the blood vessels of the gastrointestinal tract. Motility of the gastrointestinal tract is provided by two layers of smooth muscle, circular muscle and longitudinal muscle, which are interposed between the submucosa and the serosa. The longitudinal muscle layer is thin and contains few nerve fibers, whereas the circular muscle layer is thick and more densely innervated. Neurons do not make true synapses on the gastrointestinal smooth muscle fibers; rather they release transmitters from varicosities along the length of their axons. Two plexuses, the submucosal plexus and the myenteric plexus, contain the nervous system of the gastrointestinal tract. The myenteric plexus lies between the circular muscle and the longitudinal muscle. The extrinsic component is the sympathetic and parasympathetic innervation of the gastrointestinal tract. The enteric nervous system is wholly contained within the submucosal and myenteric plexuses in the wall of the gastrointestinal tract; it communicates extensively with the parasympathetic and sympathetic nervous systems. The pattern of parasympathetic innervation of the gastrointestinal tract is consistent with its function. The vagus nerve innervates the upper gastrointestinal tract including the striated muscle of the upper third of the esophagus, the wall of the stomach, the small intestine, the ascending colon, and a portion of the transverse colon. The pelvic nerve innervates the lower gastrointestinal tract including the walls of the transverse, descending, and sigmoid colons. Recall from Chapter 2 that the parasympathetic nervous system has long preganglionic fibers that synapse in ganglia in or near the target organs. In the gastrointestinal tract, these ganglia actually are located in the walls of the organs within the myenteric and submucosal plexuses. Postganglionic neurons of the parasympathetic nervous system are classified as either cholinergic or peptidergic. The vagus nerve is a mixed nerve in which 75% of the fibers are afferent and 25% are efferent. Such reflexes, in which both afferent and efferent limbs are contained in the vagus nerve, are called vagovagal reflexes. Sympathetic Innervation Preganglionic fibers of the sympathetic nervous system are relatively short and synapse in ganglia outside the gastrointestinal tract. Efferent neurons of the parasympathetic and sympathetic nervous systems synapse in the myenteric and submucosal plexuses, in the smooth muscle, and in the mucosa. Approximately 50% of the sympathetic nerve fibers are afferent and 50% are efferent. Intrinsic Innervation the intrinsic or enteric nervous system can direct all functions of the gastrointestinal tract, even in the absence of extrinsic innervation. These ganglia also receive sensory information directly from mechanoreceptors and chemoreceptors in the mucosa and send motor information directly to smooth muscle, secretory, and endocrine cells. A large number of neurochemicals, or neurocrines, have been identified in neurons of the enteric nervous system (Table 8. Some of the substances listed are classified as neurotransmitters and some are neuromodulators. Most neurons of the enteric nervous system contain more than one neurochemical, and upon stimulation, they may cosecrete two or more neurocrines. These functions include contraction and relaxation of the smooth muscle wall and the sphincters; secretion of enzymes for digestion; secretion of fluid and electrolytes; and trophic (growth) effects on the tissues of the gastrointestinal tract. Characteristics of Gastrointestinal Peptides the gastrointestinal peptides are classified as hormones, paracrines, or neurocrines. Hormones are peptides released from endocrine cells of the gastrointestinal tract. They are secreted into the portal circulation, pass through the liver, and enter the systemic circulation. The systemic circulation then delivers the hormone to target cells with receptors for that hormone. Endocrine cells of the gastrointestinal mucosa are not concentrated in glands but are single cells or groups of cells dispersed over large areas. Paracrines, like hormones, are peptides secreted by endocrine cells of the gastrointestinal tract. In contrast to hormones, however, paracrines act locally within the same tissue that secretes them. Paracrine substances reach their target cells by diffusing short distances through interstitial fluid, or they are carried short distances in capillaries. Thus for a substance to have a paracrine action, the site of secretion must be only a short distance from the site of action. The major gastrointestinal peptide with a known paracrine function is somatostatin, which has inhibitory actions throughout the gastrointestinal tract. After release, the neurocrine diffuses across the synapse and acts on its target cell. Gastrointestinal Hormones Enteroendocrine cells are specialized hormone-secreting cells of the gastrointestinal tract. Several criteria must be met for a substance to qualify as an "official" gastrointestinal hormone: (1) the substance must be secreted in response to a physiologic stimulus and be carried in the bloodstream to a distant site, where it produces a physiologic action; (2) its function must be independent of any neural activity; and (3) it must have been isolated, purified, chemically identified, and synthesized. Gastrin the functions of gastrin are coordinated to promote hydrogen ion (H+) secretion by the gastric parietal cells. Thus during the interdigestive period, most of the serum gastrin is in the G34 form, which is secreted at low-basal levels. Rather, each form of gastrin has its own biosynthetic pathway, beginning with its own precursor, a progastrin molecule.
Purchase fenofibrate overnight delivery
Testosterone caused differentiation of the wolffian ducts into the internal male genital tract (epididymis cholesterol mayo clinic cheap fenofibrate amex, vas deferens, seminal vesicles), a process that does not require dihydrotestosterone and thus occurred even though she is lacking 5-reductase. At puberty, the clitoris grew and became more like a penis because of the high-normal circulating level of testosterone; apparently, with high enough levels, the androgen receptors that mediate growth of the external genitalia can be activated. Her voice deepened and she acquired skeletal muscle mass, actions that are mediated by testosterone and do not require conversion to dihydrotestosterone. Despite acquiring many masculine characteristics, Jenny did not develop body and facial hair because the hair follicles require dihydrotestosterone. Jenny did not develop breasts because she did not have ovaries, which in normal females are the source of the estrogen required for breast development. If Jenny chooses to continue life as a woman, it will be necessary to remove her testes, which are producing the testosterone that is causing her to be selectively masculinized. In addition, because she lacks ovaries, Jenny has no endogenous source for the estrogen needed for breast development and female fat distribution; thus she would require treatment with supplemental estrogen. She may elect to have surgical correction of the introitus; however, even with the surgery, she will not be able to bear children because she lacks ovaries and an internal female genital tract. If Jenny chooses to live the rest of her life as a man, she will be treated with androgenic compounds that do not require 5-reduction for activity. The supplemental androgens will complete the masculinization process including development of male body and facial hair, sebaceous gland activity, growth of the prostate, and, in later life, male pattern baldness. The ovaries, analogous to the testes in the male, have two functions: oogenesis and secretion of the female sex steroid hormones, progesterone and estrogen. Each adult ovary is attached to the uterus by ligaments, and running through these ligaments are the ovarian arteries, veins, lymphatic vessels, and nerves. It is lined by germinal epithelium and contains all of the oocytes, each of which is enclosed in a follicle. The shortest duration for the first stage is approximately 13 years (the approximate age at first ovulation); the longest duration is 50 years (the approximate age at menopause). As the primary oocyte grows, the granulosa cells proliferate and nurture the oocyte with nutrients and steroid hormones. During this stage, the primordial follicle develops into a primary follicle, theca interna cells develop, and granulosa cells begin to secrete fluid. The second stage of follicular development occurs much more rapidly than the first stage. A single graafian follicle achieves dominance over its cohorts, and the cohorts regress. On day 14 of a 28-day menstrual cycle, ovulation occurs and the dominant follicle ruptures and releases its oocyte into the peritoneal cavity. At this time, the first meiotic division is completed and the resulting secondary oocyte enters the nearby fallopian tube, where it begins the second meiotic division. In the fallopian tube, if fertilization by a sperm occurs, the second meiotic division is completed, producing the haploid ovum with 23 chromosomes. The corpus luteum is composed primarily of granulosa cells but also of theca cells, capillaries, and fibroblasts. The corpus luteum synthesizes and secretes steroid hormones, which are necessary for implantation and maintenance of the zygote should fertilization occur. If fertilization does occur, the corpus luteum will secrete steroid hormones until the placenta assumes this role, later in pregnancy. Systemically, the ovarian steroid hormones act on a variety of target tissues including uterus, breast, and bone. The functional unit of the ovaries is the single ovarian follicle, which comprises one germ cell surrounded by endocrine cells. When fully developed, the ovarian follicle serves several critical roles: It will provide nutrients for the developing oocyte; release the oocyte at the proper time (ovulation); prepare the vagina and fallopian tubes to aid in fertilization of the egg by a sperm; prepare the lining of the uterus for implantation of the fertilized egg; and, in the event of fertilization, maintain steroid hormone production for the fetus until the placenta can assume this role. The meiotic process continues until approximately 6 months after birth, at which point all oogonia have become oocytes. The oocytes remain in a state of suspended prophase; the first meiotic division will not be completed until ovulation occurs many years later. At birth, only 2 million oocytes remain; by puberty, only 400,000 oocytes remain; by menopause (which marks the end of the reproductive period), few, if any, oocytes remain. Whereas males continuously produce spermatogonia and spermatocytes, females do not produce new oogonia and function from a declining pool of oocytes. If fertilization occurs, the corpus luteum secretes steroid hormones and supports the developing zygote. If no fertilization occurs, the corpus luteum regresses and becomes the corpus albicans. Virtually all steps in the biosynthetic pathway are the same as those discussed previously for the adrenal cortex and the testes. Recall that the adrenal cortex produces all intermediates up to the level of androstenedione, but because it lacks the enzyme 17-hydroxysteroid dehydrogenase, it does not produce testosterone. Recall also that the testes, having 17-hydroxysteroid dehydrogenase, produce testosterone as their major hormonal product. In the ovaries, all steps in the biosynthetic pathway are present including aromatase, which converts testosterone to 17-estradiol, the major ovarian estrogen. Progesterone and 17-estradiol are synthesized as follows: Theca cells synthesize and secrete progesterone. Every 28 days a sequence of follicular development, ovulation, and formation and degeneration of a corpus luteum is repeated in the menstrual cycle. The first 14 days of the menstrual cycle involve follicular development and are called the follicular phase. The last 14 days of the menstrual cycle are dominated by the corpus luteum and are called the luteal phase. At the midpoint of the cycle, between the follicular and luteal phases, ovulation occurs. In the granulosa cells, androstenedione is converted to testosterone and testosterone is then converted to 17estradiol. The follicular and luteal phases are characterized by negative feedback of estradiol and progesterone, respectively, on the anterior pituitary. Midcycle is characterized by positive feedback of estradiol on the anterior pituitary. Thus the follicular phase is dominated by negative feedback effects of estradiol on the hypothalamicpituitary axis. Estradiol levels rise sharply as a result of the proliferation of follicular cells and the stimulation of estradiol synthesis that occurred during the follicular phase. In the luteal phase of the menstrual cycle, the major hormonal secretion of the ovaries is progesterone. Thus the luteal phase is dominated by negative feedback effects of progesterone on the hypothalamic-pituitary axis. In general, the two ovarian steroid hormones function in a coordinated fashion to support reproductive activity of the female including development of the ovum, development and maintenance of the corpus luteum to sustain a fertilized ovum, maintenance of pregnancy, and preparation of the breasts for lactation. Over the course of the menstrual cycle, estrogen secretion by the ovaries precedes progesterone secretion, preparing the target tissues to respond to progesterone. An example of this "preparation" is seen in the up-regulation of progesterone receptors by estrogen in several target tissues. Without estrogen and its up-regulatory action, progesterone has little biologic activity. Conversely, progesterone down-regulates estrogen receptors in some target tissues, decreasing their responsiveness to estrogen. In turn, estrogen promotes the growth and development of the female reproductive tract: the uterus, fallopian tubes, cervix, and vagina. Progesterone is also active in these tissues, usually increasing their secretory activity.

Purchase 160 mg fenofibrate amex
The egl-1 gene inhibits the anti-apoptotic actions of ced-9 cholesterol test at walgreens purchase 160mg fenofibrate amex, thus allowing pro-apoptotic genes to become active and induce to cell death. These homologs were found to regulate pathways that are similar, though more complex. Scientists also began to explore how trophic factors interact with these pathways to regulate neuronal survival. Caspases are cysteine proteases that cleave target proteins next to an aspartate residue. Since the initial discovery of caspases, it has been determined that Caspase-9 is the homolog of ced-3. The mammalian homolog of ced-4 is Apaf-1 (apoptotic protease activating factor-1). Apaf-1 and other proteins form a complex called an apoptosome, a critical component of the cell death pathway. The homolog of ced-9 is Bcl-2, a member of a large family of molecules initially discovered as a part of the B-cell lymphoma-2 family of genes. In addition to the anti-apoptotic Bcl-2 gene, the Bcl-2 family includes multiple genes divided into two subfamilies: one anti-apoptotic, the other pro-apoptotic. The best-characterized anti-apoptotic, survival-promoting members of the Bcl-2 family are Bcl-2 and Bcl-xL, while the best-known pro-apoptotic members are Bax and Bak, which interact with several related genes to promote cell death. When the anti-apoptotic genes Bcl-2 or Bcl-xL were deleted, the number of neurons that died was greater. Conversely, when the pro-apoptotic gene Bax was deleted, there was an increase in the number of neurons that survived. It is now recognized that cleavage of several distinct pro-caspases results in the activation of the corresponding caspases that in turn mediate cell death pathways. One of the first cell death pathways described in mammals involves the release of cytochrome c from the mitochondria. When pro-apoptotic signals activate Bax or Bak, these proteins form complexes at the mitochondrial membrane that somehow induce the formation of a pore, allowing release of cytochrome c into the cytosol. Pro-caspase-9 is cleaved within the apoptosome, and the resulting mature caspase-9 functions as the initiator caspase that activates, in turn, subsequent caspases, such as caspase-3, the predominant effector caspase in neurons undergoing cell death. Trophic factors can increase neuronal survival by inhibiting the apoptotic pathway at various points. Without cytochrome c, the apoptosome does not form and the caspases that induce cell death cannot be activated. Thus, trophic factors not only stimulate survival pathways, but also actively inhibit the cell death cascade. Pro-caspase-9 is cleaved within the apoptosome, and the resulting caspase-9 cleaves pro-caspase-3. The second influential discovery regarding the extrinsic pathway of cell death was the finding that different forms of neurotrophins serve distinct roles in the developing nervous system. Neurotrophins are first generated as pro-neurotrophins-precursor molecules of approximately 270 amino acids. Pro-neurotrophins are then cleaved to yield mature neurotrophins of roughly 120 amino acids. Initially, it was thought that only the mature forms were important during neural development. In recent years, however, roles for the unprocessed, precursor forms in regulating cell death have been identified. The pro-apoptotic Bax causes cytochrome c to be released from the mitochondria, thus allowing it to form an apoptosome and activate the caspase cascade (double arrows) that leads to cell death. Other signals are also important in ensuring that the proper number of cells survive. Studies that map the pathways that regulate cell survival and death in various species and cell populations not only provide important information on how cell death occurs under normal conditions, but also generate insights that may ultimately yield interventions to halt cell death in neurodegenerative diseases. Over the long and interesting history of growth factor research, scientists have come to understand that growth factors not only promote survival by activating specific intracellular pathways, but also inhibit pathways that promote cell death. To ensure proper formation of the nervous system, all of these components must be carefully regulated to make sure the right cells are living or dying at the correct time of development. Additional roles for the neurotrophins and other growth factors continue to be discovered, and evidence now shows that such factors play multiple roles, such as regulating the expression of neurotransmitters and ion channels at various times during development and adulthood. As illustrated in many examples above, the importance of trophic factors extends beyond development, and their release is not limited to target tissues. As scientists continue to learn more about trophic factors and their expression patterns, more light is shed on their varied functions. For example, several growth factors are present in adult tissues, and some of these are up-regulated in response to injury. Growth factors are also released by different glial cells and even by neurons themselves. Hope remains that one day growth factors can be delivered effectively to regions of the nervous system that have been damaged by injury or disease to restore nerve cell function or delay neuronal death. Bothwell M (1995) Functional interactions of neurotrophins and neurotrophin receptors. Hamburger V (1939) Motor and sensory hyperplasia following limb bud transplantations in chick embryos. Levi-Montalcini R, Meyer H & Hamburger V (1954) In vitro experiments on the effects of mouse sarcomas 180 and 37 on the spinal and sympathetic ganglia of the chick embryo. Synaptic Formation and Reorganization Part I: the Neuromuscular Junction 9 T his chapter introduces two essential aspects of neurodevelopment: the process of synaptogenesis-the formation of new synaptic contacts in the nervous system-and the process of synaptic reorganization-the subsequent strengthening or loss of a subset of these connections. The study of synapses, including how they initially form and later reorganize, has a long history-one often marked by lively debates and strong differences of opinion. As noted in previous chapters, neurobiologists in the late 1800s debated whether neurons communicated through a syncytial network or through connections between individual cells. Evidence ultimately demonstrated that neurons communicated through small spaces-connections now termed synapses. In the first half of the twentieth century, scientists engaged in another debate, this one regarding whether the communication at synapses occurred primarily through chemical or electrical signals. These differing opinions were often referred to as the "war of the soups and the sparks," with the "soups" referring to chemical signals and the "sparks" to electrical signals. As scientists tested the two hypotheses, it was eventually determined that the majority of synapses utilize chemical signals in the form of neurotransmitters to mediate neural communication. Chapters 9 and 10 therefore focus on the development of structures associated with chemical synapses. While most of what is currently known about synaptic development comes from studies of the more prevalent chemical synapses, mechanisms regulating the development of electrical synapses, such as those formed by gap junctions, also remain an active area of study, and in some synaptic locations chemical and electrical signals work together to optimize neuronal communication. The establishment of functional synapses is a dynamic process that takes place over an extended period of time. For example, an extending, motile growth cone must transform into a nerve terminal capable of releasing a specific neurotransmitter, and the target cell must begin to produce the corresponding neurotransmitter receptors. In addition, numerous proteins are required to direct localized neurotransmitter release and organize receptors so they are available for rapid binding. All of these specialized proteins are also produced during the process of synaptogenesis. Perhaps surprisingly, after all of the effort to convert each synaptic partner into a highly differentiated cell, a subset of synaptic connections is lost during the normal course of development. Thus, rather than induce differentiation in only the subset of synaptic connections required for neural function, the nervous system instead over-produces highly specialized synaptic contacts, then eliminates a portion of them. The stabilization of synapses occurs in part by maintaining synapses that form functional partners. As the innervating neuron initiates action potentials in the target cell, the cells eventually begin to fire action potentials in synchrony with one another. The phrase "cells that fire together, wire together" describes one common mechanism by which cells maintain selected synaptic connections. Any target cells that do not receive sufficient input, or those that fire asynchronously with the innervating neuron, will eventually lose the synaptic connection; thus, those that are "out of sync, lose their link. This article begins with an overview of the synaptic structures found in synapses throughout the nervous system.
Generic 160 mg fenofibrate amex
Humoral hypercalcemia of malignancy is treated with furosemide cholesterol za wysoki objawy order fenofibrate 160mg with mastercard, which inhibits renal Ca2+ reabsorption and increases Ca2+ excretion, and inhibitors of bone resorption such as etidronate (Box 9. This autosomal dominant disorder is characterized by decreased urinary Ca2+ excretion and increased serum Ca2+ concentration. When the renal receptors are defective, a high serum Ca2+ concentration is incorrectly sensed as "normal" and Ca2+ reabsorption is increased (leading to decreased urinary Ca2+ [hypocalciuria] and increased serum Ca2+ concentration). It is synthesized and secreted by the parafollicular or C ("C" for calcitonin) cells of the thyroid gland. The calcitonin gene directs the synthesis of preprocalcitonin and a signal peptide is cleaved to yield procalcitonin; other peptide sequences are then removed, and the final hormone, calcitonin, is stored in secretory granules for subsequent release. The major action of calcitonin is to inhibit osteoclastic bone resorption, which decreases the plasma Ca2+ concentration. In fact, a physiologic role for calcitonin in humans is uncertain because neither thyroidectomy (with decreased calcitonin levels) nor thyroid tumors (with increased calcitonin levels) cause a derangement of Ca2+ metabolism, as would be expected if calcitonin had important regulatory functions. The man decided he did not want further treatment and would rather spend his remaining time at home with family. In the past week, the man has become lethargic and is producing large volumes of urine (polyuria). His physician admits him to the hospital, where he is found to have significantly increased serum Ca2+ concentration and decreased serum phosphate concentration. The combination of an elevated serum osmolarity and very dilute urine suggests that the man has developed diabetes insipidus, either central or nephrogenic. In the hospital, the man is given saline and furosemide (a loop diuretic) to decrease his serum Ca2+ concentration. Furosemide inhibits the Na+-K+-2Cl- cotransporter in the thick ascending limb of the kidney. When this cotransporter is inhibited, the normal lumen positive potential in the thick ascending limb is abolished, thereby inhibiting Ca2+ reabsorption and increasing Ca2+ excretion. He is sent home with a prescription for pamidronate, an inhibitor of bone resorption that will help keep his serum Ca2+ in the normal range. The role of vitamin D is to promote mineralization of new bone, and its actions are coordinated to increase both Ca2+ and phosphate concentrations in plasma so that these elements can be deposited in new bone mineral. Synthesis of Vitamin D Vitamin D (cholecalciferol) is provided in the diet and is produced in the skin from cholesterol. Vitamin D has formal "hormone" status because cholecalciferol itself is inactive and must be successively hydroxylated to an active metabolite. There are two sources of cholecalciferol in the body: It is either ingested in the diet or synthesized in the skin from 7-dehydrocholesterol in the presence of ultraviolet light. It is hydroxylated in the liver to form 25-hydroxycholecalciferol, which also is inactive. In the kidney, 25-hydroxycholecalciferol undergoes one of two routes of hydroxylation: It can be hydroxylated at the C1 position to produce 1,25dihydroxycholecalciferol, which is the physiologically active form, or it can be hydroxylated at C24 to produce 24,25-dihydroxycholecalciferol, which is inactive. Regulation of Vitamin D Synthesis Whether the renal cells produce 1,25dihydroxycholecalciferol (the active metabolite) or 24,25-dihydroxycholecalciferol (the inactive metabolite) depends on the "status" of Ca2+ in the body. When Ca2+ is insufficient, with a low dietary intake of Ca2+ and decreased plasma Ca2+ concentration, the active metabolite is preferentially synthesized to ensure that additional Ca2+ will be absorbed from the gastrointestinal tract. To increase plasma Ca2+ and phosphate concentrations, vitamin D has coordinated actions on intestine, kidney, and bone. Because 1,25-dihydroxycholecalciferol is a steroid hormone, its mechanism of action involves stimulation of gene transcription and synthesis of new proteins, which have the following physiologic actions: Intestine. The exact role of calbindin D-28K in promoting absorption in intestinal epithelial cells is uncertain. It may act as a shuttle, moving Ca2+ across the cell from lumen to blood, or it may act as a Ca2+ buffer to keep intracellular free Ca2+ low, thus maintaining the concentration gradient for Ca2+ diffusion across the luminal membrane. The actions of 1,25-dihydroxycholecalciferol on the kidney are parallel to its actions on the intestine-it stimulates both Ca2+ and phosphate reabsorption. This action may seem paradoxical because the overall action of 1,25dihydroxycholecalciferol is to promote bone mineralization. Signal peptides and other peptide sequences are cleaved from preprohormones to form the peptide hormones, which are packaged in secretory granules. Steroid hormones are synthesized from cholesterol in the adrenal cortex, testes, ovaries, and placenta. Hormone synthesis and secretion are regulated by negative and positive feedback mechanisms. Hormone receptors are also regulated by increasing (up-regulation) or decreasing (down-regulation) their number or activity. The connection between the hypothalamus and the posterior lobe of the pituitary is neuronal. The cell bodies are in the hypothalamus, and the hormones are secreted from nerve terminals in the posterior lobe of the pituitary. The hypothalamus is connected to the anterior lobe of the pituitary by hypothalamichypophysial portal blood vessels. Growth hormone is required for growth to normal stature and has actions on carbohydrate metabolism, protein synthesis, organ growth, and bone growth. Prolactin secretion is under tonic inhibition, mediated by dopamine from the hypothalamus. This condition is rare in areas of the world where vitamin D is supplemented and when there is adequate exposure to sunlight. In adults, vitamin D deficiency results in osteomalacia, in which new bone fails to mineralize, resulting in bending and softening of the weight-bearing bones. Vitamin D resistance occurs when the kidney is unable to produce the active metabolite, 1,25dihydroxycholecalciferol. This condition is called "resistant" because no matter how much vitamin D is supplemented in the diet, it will be inactive because the C1 hydroxylation step in the kidney is absent or is inhibited. Vitamin D resistance can be caused by the congenital absence of 1-hydroxylase or, more commonly, by chronic renal failure. Chronic renal failure is associated with a constellation of bone abnormalities including osteomalacia, a consequence of the inability of the diseased renal tissue to produce 1,25-dihydroxycholecalciferol, the active form of vitamin D. Oxytocin secretion is stimulated by suckling and is responsible for milk ejection from the lactating breast. Adrenocortical steroid hormones are glucocorticoids, mineralocorticoids, and adrenal androgens, all of which are synthesized from cholesterol. Glucocorticoids stimulate gluconeogenesis and have antiinflammatory and immunosuppressive actions. Mineralocorticoids stimulate Na+ reabsorption and K+ and H+ secretion by the kidney. The islets of Langerhans have three cell types:, which secrete glucagon;, which secrete insulin; and, which secrete somatostatin. Insulin is the hormone of "abundance" and promotes storage of glucose as glycogen, storage of fatty acids in adipose, and storage of amino acids as protein. Glucagon is the hormone of "starvation" and promotes utilization of stored nutrients. Vitamin D is converted to its active form, 1,25-dihydroxycholecalciferol, in the kidney. Its actions are to increase intestinal and renal Ca2+ and phosphate absorption and to increase bone resorption. Challenge Yourself Each numbered question begins with an endocrine disorder or a disturbance to an endocrine system. The male gonads, the testes, are responsible for development and maturation of sperm and synthesis and secretion of the male sex steroid hormone, testosterone. The female gonads, the ovaries, are responsible for development and maturation of ova and synthesis and secretion of the female sex steroid hormones, estrogen and progesterone. During the first 5 weeks of gestational life, the gonads are indifferent or bipotential-they are neither male nor female. Therefore, genetic sex normally determines gonadal sex, and the gonads appear in males slightly before they appear in females.
