

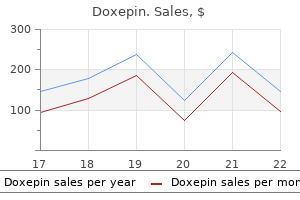
Order doxepin 75 mg free shipping
To then realize that the divergence of lymphocytes into two lineages occurred in the ancestor of jawed and jawless fish was astonishing anxiety definition discount doxepin 25 mg on-line. Now to have uncovered a primitive type of thymus candidate in jawless fish is a total shock. The genome projects in cnidarians demonstrated that certain gene families were more ancient than previously believed (from studies of the classical models Drosophila and C. The third genomewide duplication (3R) is believed to have been a major force in this instability and rapid evolution. Amphibians with metamorphosis offer models to study two modalities of self-tolerance, metamorphosis, and polyploidy. The larval immune system must somehow be suppressed with the appearance of adult self-determinants during metamorphosis and the refurbishing of the immune system. The amphibian Xenopus speciates by polyploidy, allowing an experimental system to examine the effects of whole-genome duplications on immune system loci (silencing, deletion, gene conversion, conservation), a key issue given the importance attributed to the genomewide duplications in the evolution of immune systems in vertebrates. The genomewide duplications early in the evolution of the vertebrates (the 2R hypothesis) have been confirmed in a variety of studies, and they were indeed important in the Big Bang emergence of the adaptive immune system. With the genome projects and advances in molecular biology, we have made strides in understanding old problems in vertebrate adaptive immunity, such as 1) when the L chains emerged and what the significance of more than one isotype is; 2) which antigen receptor (if any of the extant ones) came first, and all of them came to evolve to their present state; 3) when IgD emerged and what its function is in different vertebrate phyla, especially the transmembrane form; and 4) how / T cells recognize antigen. We propose that are two arms of the / T-cell lineage, one innate and the other adaptive, similar to B cells and / T cells. The long-awaited molecular mechanisms of histocompatibility in nonvertebrates have been at least partially uncovered in plants, Botryllus, and Hydractinia. However, there are tantalizing links to immune gene clusters found throughout the animal kingdom that suggest there may be an underlying fundamental similarity in recognition, perhaps related to 1) the paucity of surface receptors available because of multiple constraints resulting in sharing of receptor/ligands, and 2) the signaling cascades. Despite the great variation in mechanisms of allorecognition across phyla, are there commonalities Could this region be the conserved and link aspects of immunity between vertebrates and invertebrates The comparison of allorecogniton in invertebrates has perhaps helped to focus our attention on genetic regions that may have provided the "context" for leukocyte interactions. These effectors can activate and fix complement and they can bind to Fe receptors on the surfaces of granulocytes, monocytes, platelets, and other components of the immune response. Both activation of complement and binding to Fe receptors can contribute to the induction or maintenance of inflammation. They also provide the immune system with a polyclo al set of diverse ligand binding sites, which allow Igs, as a population, to recognize an almost unlimited array of self- and non-self-antigens, which may range from compolJllas as fundamental to life as deoxyribonucleic aci to manmade molecules that could not have played a role in the evolution of the immune system. Differential splicing allows individual lg molecules to serve as either membrane-bound receptors for the B cell that allow antigen-specific activation or as soluble effectors, which act at a distance. In vivo, proper effector function requires more than just antigen-specific binding; it requires successful neutralization of the offending antigen while avoiding potentially pathogenic self-reactivity. The receptor and effector functions of each individual lg can be localized to a separate region or domain of the molecule. Each variable (V) or constant (C) domain consists of approximately 110 to 130 amino acids, averaging 12,000 to 13,000 kD. A typical light (L) chain will thus mass approximately 25 kD, and a three C domain Cy H chain with its hinge will mass approximately 55 kD. Each of the chains contains single amino-terminal V lg domain and one, tbree, or four carboxy-terminal C lg domains. Jsion of regions of impressive sequence variability with regions of equally impressive sequence conservation. The V domains demonstrate the greatest molecular heterogeneity, with some regions including non-germline-encoded variability and others exhibiting extensive germline conservation across 500 million years of evolution. Thus it is the V domains that encode the receptor function and define the monovalent specificity of the antibody. The germline exonic derivation of the sequence is shown at the top, and the protein structure is shown at the bottom. The location of the various cysteine residues that help hold both the individual domains and the various Ig subunits together are illustrated. Papain digests IgG molecules above the cysteine residues in the hinge that holds the two H chains together yielding two Fab molecules and an Fc, whereas pepsin digests below releasing an (Fab)2 fragment and two individual Fcs (which are typically degraded to smaller peptide fragments). In their role as antibodies, Igs are released into the circulation from where they may traffic into the tissues and across mucosal surfaces. In their role as the B-cell antigen receptor, they are anchored to the membrane by means of their M1:M2 transmembrane domain. Soluble antibodies can also be pressed into service as heterologous cell surface antigen receptors by means of their attachment to membrane-bound Fc receptors. The major difference between these two forms of cell surface receptors is that Igs as B-cell antigen receptors provide a monoclonal receptor for each B cell, whereas antibodies bound to Fc receptors endow the cell with a polyclonal set of antigen recognition molecules. Paratopes and Epitopes the immunoglobulin-antigen interaction takes place between the paratope, the site on the Ig at which the antigen binds, and the epitope, which is the site on the antigen that is bound. It is important to appreciate that antibodies do not recognize antigens; they recognize epitopes borne on antigens. It also is one scenario that permits the same antibody to bind divergent antigens that share equivalent epitopes, a phenomenon referred to as cross-reactivity. It has been estimated that triggering of effector functions in solution typically requires aggregation of three or more effector domains, and thus tends to involve the binding of three or more epitopes. For antigens encoding diverse epitopes, which is more typical of monodisperse single-domain molecules in solution, triggering of inflammatory effector functions may require the binding of a diverse Isotypes and Idiotypes Igs can also serve as antigens for other Igs. The heavy-chain constant regions (green) also include the hinge (yellow) between the first two domains. The heavy- and light-chain variable regions (red and dark blue, respectively) are N terminal to the heavy- (green) and light-chain (light blue) constant regions. Complementarity determining region loops in the heavy- and light-chain variable regions (yellow and white) are illustrated as well. Recognition of these isotypes first allowed grouping of Igs into recognized classes. Each class of Ig defines an individual set of C domains that corresponds to a single H chain constant region gene. Examples include the cross-reactive idiotypes found on monoclonal IgM rheumatoid factors derived from individuals with mixed cryoglobulinemia, each of which can be linked to the use of individual V gene segments. This section describes the history of the identification of Ig and introduces fundamental terminology. Antibodies and Antigens Aristotle and his contemporaries attributed disease to an imbalance of the four vital humors: the blood, the phlegm, and the yellow and black biles. The Fab arms of the antibody targets and probes are drawn to indicate their rotational orientation as planar (oval with open center), intermediate (bone shape with or without central opening), or perpendicular ("dumbbell shaped"). Different complexes illustrate the range of Fab-Fab angles made possible by segmental flexibility. Antibody complexes were stained with 2% uranyl formate as described by Roux et al. Gamma Globulins In 1939, Tiselius and Kabat immunized rabbits with ovalbumin and fractionated the immune serum by electrophoresis into albumin, alpha-goblulin, beta-globulin, and gamma-globulin fractions. Immunoelectrophoresis subsequently permitted identification of the various Ig classes and subclasses. Fab and Fc In 1949, Porter first used papain to digest IgG molecules into two types of fragments, termed Fab and Fc (Table 5. The structure of the -pleated sheets in an Ig domain varies depending on the number and conformation of strands in each sheet. C-type domains, which are the most compact, have seven antiparallel strands distributed as three strands in the fi rst sheet and four strands in the second. Each of these strands has been given an alphabetical designation ranging from amino terminal A to carboxy-terminal G. Side chains positioned to lie sandwiched between the two strands tend to be nonpolar in nature. This hydrophobic core helps maintain the stability of the structure to the point that V domains engineered to replace the conserved cysteines with serine residues retain their ability to bind antigen.
Order doxepin
However anxiety disorder symptoms dsm 5 purchase doxepin with paypal, most pathogens enter the host through mucosal epithelia; thus, an ideal vaccine should induce protective immune responses in these tissues. Dimeric secretory IgA (sIgA) is the predominant Ig found in respiratory and gastrointestinal tract mucosal secretions, saliva, and breast milk. Although all these routes are amenable to the application of vaccines, in practical terms, only nasal and oral administrations are suitable options for all ages, genders, and cultures. Intranasal administration of vaccines by spraying, nebulization, or aerosolization induces sIgA in the airway mucosa and in the genital tract but is rather inefficient at generating intestinal antibody responses. However, intranasal vaccine delivery has raised safety concerns after reports of Bell palsy adverse effects associated with retrograde passage of inhaled antigens or adjuvants through the olfactory epithelium. To date, the only licensed vaccine administered intranasally is a live-attenuated influenza vaccine. Conversely, the ingestion of antigens (orogastric route) can induce sIgA responses in the small intestine, proximal colon, and mammary and salivary glands but is poorly efficient for the distal segments of the gut and the respiratory and reproductive tracts. The major obstacle to transcutaneous immunization, however, resides in the stratum corneum, the outermost layer of the skin that provides a natural barrier against foreign materials. Gentle disruption of this outer layer is necessary to improve vaccine delivery in the skin epidermis and requires the usage of special devices. Sublingual and transcutaneous routes of delivery of vaccines are alternatives that bypass the need for needles and represent promising new venues. Viral Vaccines Poliomyelitis Development of a vaccine against poliomyelitis represents an important achievement in the history of vaccines. It demonstrated that two different approaches, based on inactivated or live-attenuated formulations, could be successfully applied to prevention of this disabling disease. Although generally associated with the names of Salk and Sabin, discovery of the polio vaccine benefited from several key findings that preceded their work. These were the isolation of the poliovirus in monkeys, the identification of three immunologic different serotypes of the virus, the demonstration that neutralizing antibodies against one serotype could not protect against the other two, and finally the propagation of the virus in vitro in human embryonic cells of nonneural origin. Indeed, early efforts in the 1930s to obtain a vaccine using inactivated poliovirus isolated from monkey spinal cord were unsuccessful, with induction of the disease at an unacceptable rate of 1 in 1000 vaccinated individuals. This vaccine was 80% to 90% efficacious in one of the biggest field trials in the history of vaccines,159 and its introduction in the United States resulted in a dramatic decline in the cases of poliomyelitis in few years. The infections were caused by two lots of vaccine produced by Cutter Laboratories, in which the virus was not adequately inactivated, despite having passed safety controls. The presence of neutralizing antibodies at levels above a 1:8 dilution in the serum of vaccinated subjects is considered a correlate of protection. The efficacy of this vaccine depends on the capacity of the virus to infect and multiply in the gut of the vaccinated individuals. The incidence of vaccineassociated paralytic poliomyelitis can be estimated in one case per 1. It is broadly diffuse around the world with epidemic outbreaks associated with the winter season in temperate climate regions, whereas it has no seasonal pattern in tropical regions. Although often asymptomatic or self-limited, seasonal influenza infection can be debilitating, resulting in the loss of many working days every year. Moreover, it can be associated with life-threatening complications, especially in some at-risk populations such as the very young, the elderly, and the chronically ill. Annual seasonal influenza epidemics result in about three to five million cases of severe illness and about 250,000 to 500,000 deaths worldwide. Periodically, the human population is affected by influenza pandemics, characterized by rapid global spread of this infection and an increase in morbidity and mortality compared to seasonal influenza. Influenza types B and C viruses show low variability and infect mainly humans, but only virus B is responsible of epidemic episodes. On the other hand, type A virus can also infect mammals other than humans, such as pigs, dogs, and horses, and domestic or wild birds. Point mutations are continuously introduced during influenza virus replication because the enzymes that replicate the viral genome have no proofreading activity. These mutations, which are called "antigenic drifts," are responsible throughout the years of small changes in the antigens of the circulating viruses and can be observed in both types A and B viruses. These changes confer a selective advantage to the new virus, against which large segments of the population have insufficient immunity, requiring the preparation of a new vaccine every year to immunize the susceptible populations. Following the recommendations of this surveillance system, yearly vaccines for active prophylaxis of influenza are produced ad hoc and distributed worldwide to protect against the new variants of the influenza virus that are predicted to be the most widely circulating. A second type of variations that is observed only in influenza virus A is called "antigenic shifts. This reassortment can take place when both avian-specific and human-specific subtypes infect pigs, which express both sialic acid 2,3 and 2,6 galactose and therefore are susceptible to both viruses. The exchange of genetic material can lead to the formation of a new subtype that is able to infect humans and will be very different from the previously circulating viruses. As no immunologic memory for this virus exists in the human population, the new virus will rapidly spread worldwide resulting in a global pandemic outbreak. The other two pandemics which occurred in the 20th century were the Asian flu in 1957 and the Hong Kong flu in 1968, which were caused by H2N2 and H3N2 viruses, respectively. In 1977, there was a reemergence of a H1N1 virus which, however, had a limited spread and did not cause a real pandemic but only several outbreaks. However, this H1N1 virus continued to circulate and therefore was added as a third strain in the yearly vaccine formulation. Indeed, H3N2 subtype was still responsible of seasonal influenza A outbreaks from 1968 to 2009 and therefore was kept in the yearly vaccine composition. More recently, in April 2009, a new strain of H1N1 was isolated in Mexico and California and was associated with severe to fatal cases of flu. The new virus derived from the reassortment of a previous triple reassortment of bird, swine, and human flu viruses, which was circulating among pigs in North America, with a Eurasian pig flu virus; it was called "swine flu. Vaccine manufacturers initiated efforts to develop vaccines against the pandemic strain, and different adjuvanted and nonadjuvanted monovalent pandemic vaccines were licensed and used in national vaccination campaigns. Alternatively, a pandemic influenza can originate when an avian virus infects humans and can be easily transmitted from person to person. This is the fear of what might happen with a highly pathogenic strain of bird influenza, the A/H5N1 virus, which was isolated in humans for the first time in 1997 in Hong Kong. The infection has spread to wild birds and chickens in many countries in Asia, Africa, and Europe, directly causing the death of tens of million of birds and requiring the sacrifice of hundreds of million of domestic chickens in the attempt to contain spreading of the infection. Although no transmission from human to human has been observed, the H5N1 virus has the potential of originating a new severe influenza pandemic. The first vaccine against influenza was made available to the public in 1945 and was a formalin-inactivated whole virus vaccine from viruses grown in embryonated hen eggs. Most vaccines currently in use is still produced from virus cultured in eggs and then inactivated; however, additional steps in the production process have been added to obtain more purified viral preparations. A detergent treatment that eliminates the viral envelope and other purification steps are used in the production of "split" influenza vaccines, which contain only some viral components. Even cleaner vaccine preparations, called "subunits," are obtained by additional purification procedures and mainly contain the viral surface proteins. Nonetheless, two main reasons have convinced vaccine manufacturers to search for an alternative to eggs for growing influenza virus for vaccine production. First, the spreading of an avian infection caused by flu or other viruses could greatly reduce the availability of hens and thus the eggs required for vaccine manufacturing. Second, each new influenza virus has to be adapted to grow in eggs with the risk that antigenic mutations will rise in the egggrown virus compared to the virus that is circulating in the human population, ultimately negatively affecting the vaccine efficacy. Not all influenza virus strains grow well in eggs, and this can affect yield of vaccine production. In the 1990s, the circulating B strain diverged into two separate subtypes, which in the last two decades circulated with a different prevalence during each season, but the vaccine contains only the strain that will most likely circulate. However, during some seasons, the vaccine strain did not match the most widely circulating strain, resulting in reduced efficacy for the vaccine, especially in children who are the population most affected by influenza B virus. This adjuvanted vaccine shows increased immunogenicity and better clinical protection compared with the split conventional influenza vaccine in the elderly population. However, none of these strains was able to give a really attenuated strain to be used as vaccine. A live-attenuated influenza vaccine can therefore be obtained each year from two cold-adapted strains of A and B subtypes by reassortment with the seasonal wild-type viruses. This vaccine contains three attenuated reassortant strains that express the antigens from the seasonal recommended strains and are grown in pathogen-free eggs.
Cheap doxepin 10 mg on-line
For example anxiety young child discount 25 mg doxepin, rapamycin inhibits the mammalian target of rapamycin pathway, which is important for cell division and can induce anergy in T cells. Forced upregulation of p27kip induces an anergy-like state, and accordingly anergy cannot be induced in p27kip-deficient cells. Breaking T-Cell Tolerance Autoimmunity can arise by many different mechanisms, some of which include the failure of tolerance mechanisms described previously. There is much interest in uncovering strategies to break tolerance, either to promote antitumor immunity or limit autoimmunity. Correlative studies have been reported by many groups that demonstrate that patients with a defi ned T-cell signature have a better prognosis. Initially, these transgenes were introduced in multiple copies cointegrated at a random chromosomal locus. Subsequently, the transgenes were targeted as single copies integrated into the endogenous IgH and IgL chromosomal loci. Collectively, these models have revealed that actively acquired tolerance in B cells is extensive. These foreign-specific B cells matured and recirculated amongst peripheral lymphoid tissue and, when mice were immunized with the foreign hen egg lysozyme antigen, these B cells were efficiently stimulated into clonal proliferation and antibody formation. Parallel sets of transgenic mice engineered to express hen egg lysozyme as a "neo-self" protein displayed actively acquired tolerance and were specifically unable to make antibody upon immunization with lysozyme. B: Members of the B7 family are expressed on a variety of cells other than the hematopoietic system, such as the islets cells in the pancreas. It is possible that the B7 family of molecules contribute to tissue homeostasis in a variety of settings. Self-reactive B cells remained "ignorant" and exhibited little functional or numerical change when they developed in mice with 10 -10 M circulating antigen in their blood, which was only sufficient to engage a few percent of the lysozyme binding receptors on the B cells. For simplicity, this state is referred to as B cell clonal anergy but, as with T cell clonal anergy discussed previously, this single term should not be mistaken to connote a single mechanism. In fact, there are multiple separate inhibitory processes responsible for actively acquired tolerance in "anergic" B cells that are imperfectly understood, and these appear to vary depending on the nature of the autoantigen and antibody. Anergic B cells encountering monomeric lysozyme were similarly arrested as a shortlived IgMlow IgDlow immature population in the spleen in mice that contained normal numbers of competing B cells. Flow cytometric plots are gated on spleen B cells, with each dot representing one cell. These show on the x-axis the log10 relative abundance of surface immunoglobulin (Ig) on each B cell, stained with antibody to kappa light chain. In mice inheriting IgH and IgL transgenes, almost all the B cells bind lysozyme monospecifically, and cell-to-cell variation in binding is tightly correlated with variation in the amount of surface Ig on each cell. Clonal ignorance is observed in double-transgenic mice expressing the lysozyme gene selectively in the thyroid epithelium: the frequency of lysozyme binding cells is unaltered and the amount of surface Ig they display is decreased by less than twofold. In double transgenic mice where monomeric lysozyme accumulates to higher concentrations in the circulation and continuously engages 50% of the antigen receptors on the developing B cells, actively acquired tolerance in the B cells is reflected by selective downregulation of their surface IgM receptors but the cells mature to the IgDhigh recirculating stage. In a matched double transgenic strain where the same proportion of receptors is engaged by dimeric lysozyme, IgM and IgD surface antigen receptors are downregulated accompanied by developmental arrest at the T1-T2 stage, and edited B cells accumulate with normal densities of antigen receptors that do not bind lysozyme. Finally, in a double-transgenic strain where lysozyme is displayed in polyvalent form on the membrane of other blood and marrow stromal cells, lysozyme-binding B cells are almost completely eliminated and replaced by edited B cells. Green means "go"; yellow means moderate warning (moderately self-reactive); orange means strong warning (strongly self-reactive); and red means "stop" (very self-reactive). Monospecificity, Polyspecificity, and the Need to Balance Immunity with Self-Tolerance An overwhelming lesson from the studies summarized in Table 32. Why are so many self-reactive B cells allowed to reach the spleen and circulation There are compelling mathematical arguments that germline antibodies need to be polyspecific to provide adequate coverage of microbial epitopes with a fi nite pool of preimmune B cells. Increasing the crossreactivity and polyspecificity of each antibody minimizes the chance of having a gap in the repertoire due to limited B cells but this is offset by progressively larger holes in the antibody repertoire due to these antibodies also binding self-antigens. Self-reactive B cells are eliminated in a series of steps that balances the need for self-tolerance against the need to maintain clones for immunity. The degree of autoreactivity is a function of the amount of autoantigen presented and the avidity with which it is bound by the clone. Most newly formed B-cell clones have low but appreciable self-reactivity, but only a subset appear to exceed an inherited set point and trigger either elimination in the bone marrow or editing to lower self-reactivity. Clones with less autoreactivity are exported to the periphery, illustrated by the hypothetical distribution of clones in the middle. By this time, repeated binding of autoantigens has tuned down surface immunoglobulin signaling in the more self-reactive end of the spectrum, illustrated by orange shading. Binding of autoantigen and competition for follicular niches also trigger exclusion and death of the more self-reactive clones in the T-cell zones, although these cells can potentially be rescued by T cells if they bind foreign antigens with much higher avidity than they bind to self. As a result of follicular competition, together with hypermutation and further selection in germinal centers, the B-cell repertoire that recirculates for weeks or months among lymphoid tissues is skewed toward a small subset of B cells with the least autoreactivity (bottom). A much larger range of newly produced clones is nevertheless available in the T-cell zones of the spleen (middle) to be tested for its fit against microbial antigens and potentially recruited for transient antibody responses or remodeling in germinal centers. Nemazee413 and Weigert414 independently recognized that the transgenic mice had illuminated an entirely new mechanism for actively acquired tolerance: instead of clonal selection as envisaged by Burnet, this was a system for receptor selection. In the transgenic experiments, receptor editing was nevertheless inefficient because the transgenic light chain gene was located at a random chromosomal site where it was not flanked by upstream V elements that could be joined to downstream J elements. Indeed, when competitive bone marrow chimeras were established, bone marrow B cells that started out with prerearranged but self-reactive receptors nevertheless produced edited mature B cells as efficiently as competing B cells that needed to rearrange their H and L chain genes de novo. Downregulation of surface IgM by self-antigen allows Foxo1 to accumulate, reactivate Rag1 and Rag2, and induce a second round of light chain recombination. Anergy in Recirculating Immunoglobulin Dhigh B Cells the longer-lived, recirculating self-reactive B cells are characterized by selective downregulation of surface IgM but high surface IgD. Selective downregulation of IgM occurs in the absence of antigen-binding when inositol-3-phosphate activity is not dampened by the enzyme inositol 1,4,5-trisphosphate 3-kinase B. Anergic, self-reactive B cells with this phenotype represent 5% of the peripheral B cell pool in normal mice. In other words, they are not unresponsive but have tuned down their responsiveness to antigen, similar to that suggested for selfreactive T cells (see previous discussion). This is best exemplified by the short-lived anergic B cells that result from an encounter with the H-2K b protein in the liver while en route to the spleen. Reversal from anergy in this instance may result from the absence of liver-specific antigen in the spleen. In Ig-transgenic mice expressing an antibody against erythrocytes, expansion of B1 cells depends upon their relative shielding from erythrocytes in this compartment476,477 and requires concurrent infection with pathogenic or commensal microorganisms. In either case, the IgM antibody that is secreted binds with high avidity to erythrocytes and causes severe autoimmune hemolytic anemia. A similar situation occurs in Ig-transgenic mice expressing a low-affinity germline antibody against a carbohydrate epitope on the T-cell protein, Thy1, where the B cells are arrested as anergic immature cells in the spleen, few reach the lymph nodes, but they accumulate as B1 cells in the peritoneal cavity and secrete anti-Thy1 IgM autoantibody. Remarkably, the induction of B1 cells and autoantibodies requires the Thy1 autoantigen and is abolished in Thy1-deficient mice. It is not yet known if this also requires infection with commensal or pathogenic microbes, like the antierythrocyte B1 cells. When Thy1 is restored at 10% of normal levels in the mice, it changes the fate of the self-reactive B cells again by inducing their maturation into marginal zone B cells that do not secrete autoantibody. But it is fair to say that we are only in the early parts of a play of three acts. The first act has been to define the physiologic mechanisms of actively acquired tolerance. There are likely to be entirely new mechanisms revealed in the future, and answers are yet to come about fundamental questions like how lymphocytes choose alternative fates of death, survival, or growth at a given tolerance checkpoint. Currently, this is an area of great activity, through genome-wide association studies of unparalleled size and the dawn of an exciting era of whole exome and genome sequencing in patient cohorts.
Buy 75 mg doxepin mastercard
Some of these disorders are caused by mutations in genes that are broadly expressed and others are caused by defects affecting hematopoietic cells alone but involving processes not specific to immune defense anxiety group therapy purchase doxepin 75mg line. Interestingly, defects in the same signal transduction pathway or biologic process may cause different clinical manifestations. Another two immunodeficiencies resulting in abnormal regulation of the actin cytoskeleton have recently been described. A single patient has been reported with microcephaly and hypomorphic mutations of the Rad50 gene. Patients with these conditions display selective susceptibility to recurrent, invasive Neisseria infections. Remarkably, alternative and terminal complement defects are almost exclusively associated with gonorrheal and meningococcal diseases. Schematic diagram of cytokine production and cooperation between phagocytes/dendritic myeloid cells and natural killer/T lymphocytes. Surprisingly, however, these patients are not prone to infections with other intracellular agents, including most viruses, in particular. The rare viral illnesses occurring in these patients may have been favored by mycobacterially induced immunosuppression. Finally, these studies lend weight to the idea that otherwise healthy children with other infectious diseases may suffer from single-gene inborn errors of immunity. It is common in patients with various inherited or acquired T-cell deficits that are also associated with other infections. The only consistent immunologic abnormality is a lack of antibodies directed against glycans. Gram-negative infections are rare, with the exception of those caused by Pseudomonas and, more rarely, Shigella. Surprisingly, these patients are susceptible to a few pyogenic bacteria, but normally resistant to other bacteria and parasites, fungi, and viruses. The clinical status of patients improves spontaneously from adolescence onwards, with no deaths or invasive infections observed in the absence of prophylaxis. Proteins for which genetic mutations have been identified but not associated with susceptibility to infectious diseases are shown in red. Tonal signaling in response to self-antigens is essential to promote the progression of T- and B-cell development in the thymus and bone marrow, respectively. However, the prevention of autoimmunity requires T- and B-lymphocytes with high affinity for self-antigens to be deleted or kept in check. In addition, some rare monogenic disorders associated mostly, if not exclusively, with autoimmunity highlight the critical role played by tolerogenic mechanisms in immune homeostasis and function (see Table 48. The autoimmune phenotype of autoimmune polyendocrinopathy type I patients might well be both broader and more profound in the absence of neutralization of these cytokines, which have been shown to play an important role in autoimmunity in the mouse model. Treg cells may be generated in the thymus (natural Treg) or in the periphery (induced Treg), and these cells have suppressive activity. Inborn Errors of Apoptosis A syndrome consisting of generalized lymphadenopathy, autoimmune cytopenia, and hypergammaglobulinemia was described in 1967 by Canale and Smith. Deep sequencing of the exomes and genomes of patients with unexplained autoimmunity will undoubtedly shed new light on the mechanisms governing central and peripheral tolerance in B cells and T cells. Postmortem examination revealed a prominent infi ltrate of lymphocytes, plasma cells, and histiocytes in the liver, spleen, and bone marrow. In some cases, the disease occurred in multiple family members and was thought to be intrinsic, whereas the sporadic presentation in other patients was thought to be a consequence of infections or cancers. This led to a distinction being made between primary and secondary forms of hemophagocytic lymphohistocytosis, although it is now clear that many sporadic cases are actually genetic in origin and that episodes of familial cases can be triggered by infection (Table 48. The study of patients with hemophagocytic lymphohistiocytosis has made an important contribution to definition of the role of lymphocyte cytotoxicity in immune homeostasis. Upon recognition of target cells, cytotoxic T-lymphocytes mobilize cytolytic granules toward the point of contact with the target cell. The Rab27a protein mediates docking of the cytolytic granule; the syntaxin-11/Munc18-2 complex also participates at this process. Then, Munc13-4 favors priming of the granule, allowing a shift of syntaxin-11 from a closed to an open conformation. Eventually, the membrane of the cytolytic granule fuses with the cell membrane and granzyme, contained in the cytolytic granules is delivered to the target cell through pores formed by perforin. Optimal results have recently been obtained with reduced-intensity conditioning regimens. At the postmortem examination, lymphocytic and histiocytic infi ltration was apparent in multiple organs. The field is so large and diverse that we could easily have selected other topics. For example, we did not review the inherited disorders of complement, despite their recently discovered surprising association with hemolytic uremic syndrome and related disorders. As host defenses are mediated not only by leukocytes, but by almost all cells and tissues, a myriad of diseases, infectious and otherwise, may result from genetic lesions affecting host defense genes. In any case, we have attempted to highlight the main clinical and immunologic implications of these studies. Over the last 60 years, more than 200 disorders have been clinically described, immunologically deciphered, and genetically dissected. This field has, arguably, been one of the most successful in pediatrics and clinical immunology. The immunologic lessons learned from these experiments of nature are also of considerable interest. Indeed, whether identified by genome-wide or candidate gene approaches, these genes have, in some cases, been assigned a new function, or at least had their known function redefined, in the human model. There are grounds for optimism, as the increasingly careful and widespread clinical care of 7 billion patients will provide an extraordinary wealth of phenotypic description. Moreover, spectacular technologic progress in genetics is making it possible to study the genome of these patients rapidly, searching for morbid lesions at high speed. The identification of causal relationships between gene lesions and clinical phenotypes is based on the molecular and cellular dissection of immunologic pathogenesis. These studies will be of benefit to patients, while providing new insight into the function of host defense genes. The seeds of disease: an explanation of contagion and infection from the Greeks to the Renaissance. The discovery of microorganisms by Robert Hooke and Antoni Van Leeuwenhoek, fellows of the Royal Society. Micrographia: Or Some Physiological Descriptions of Minute Bodies made by Magnifying Glasses with Observations and Inquiries thereupon. Microscopic Investigations on the Accordance in the Structure and Growth of Plants and Animals. Membranes: Metaphors of Investion in Nineteenth Century Literature, Science and Politics. A Linnaean thesis concerning Contagium Vivum: the "Exanthemata Viva" of John Nyander and its placer in contemporary thought. The effect of prolonged cultivation in vitro upon the pathogenicity of yellow fever virus. Translatonal mini-review series on vaccines: the Edward Jenner Museum and the history of vaccination. An experimental investigation of the role of the blood fluids in connection with phagocytosis.
Buy doxepin 10 mg low price
Structure of the quaternary complex of interleukin-2 with its alpha anxiety symptoms tingling order doxepin with a mastercard, beta, and gammac receptors. Critical cytoplasmic region of the interleukin 6 signal transducer gp130 is conserved in the cytokine receptor family. Convergent mechanisms for recognition of divergent cytokines by the shared signaling receptor gp130. Molecular and structural basis of cytokine receptor pleiotropy in the interleukin-4/13 system. Interleukin-2 receptor gamma chain: a functional component of the interleukin-7 receptor. Interleukin-2 receptor gamma chain: a functional component of the interleukin-4 receptor. The molecular basis of X-linked severe combined immunodeficiency: defective cytokine receptor signaling. Gene for T-cell growth factor: location on human chromosome 4q and feline chromosome B1. A monoclonal antibody that appears to recognize the receptor for human T-cell growth factor; partial characterization of the receptor. Novel interleukin-2 receptor subunit detected by cross-linking under highaffinity conditions. Demonstration of a non-Tac peptide that binds interleukin 2: a potential participant in a multichain interleukin 2 receptor complex. Interleukin 2 high-affinity receptor expression requires two distinct binding proteins. Interleukin-2 receptor signaling: at the interface between tolerance and immunity. Central role of defective interleukin-2 production in the triggering of islet autoimmune destruction. Mast cell interleukin-2 production contributes to suppression of chronic allergic dermatitis. Priming for T helper type 2 differentiation by interleukin 2-mediated induction of interleukin 4 receptor alpha-chain expression. Type 1 and type 2 cytokine dysregulation in human infectious, neoplastic, and inflammatory diseases. Janus-kinase-3-dependent signals induce chromatin remodeling at the Ifng locus during T helper 1 cell differentiation. Hematopoietin sub-family classification based on size, gene organization and sequence homology. The murine interleukin-4 receptor: molecular cloning and characterization of secreted and membrane bound forms. Human interleukin 4 receptor confers biological responsiveness and defines a novel receptor superfamily. Recombinant interleukin 7, pre-B cell growth factor, has costimulatory activity on purified mature T cells. The defective gene in X-linked severe combined immunodeficiency encodes a shared interleukin receptor subunit: implications for cytokine pleiotropy and redundancy. Cloning of the human and murine interleukin-7 receptors: demonstration of a soluble form and homology to a new receptor superfamily. Expression of interleukin 9 in the lungs of transgenic mice causes airway inflammation, mast cell hyperplasia, and bronchial hyperresponsiveness. The absence of interleukin 9 does not affect the development of allergeninduced pulmonary inflammation nor airway hyperreactivity. Interferon-regulatory factor 4 is essential for the developmental program of T helper 9 cells. A lymphokine, provisionally designated interleukin T and produced by a human adult T-cell leukemia line, stimulates T-cell proliferation and the induction of lymphokineactivated killer cells. The biology of interleukin-2 and interleukin-15: implications for cancer therapy and vaccine design. Interleukin 15 controls the generation of the restricted T cell receptor repertoire of gamma delta intestinal intraepithelial lymphocytes. Interleukin 15-dependent crosstalk between conventional and plasmacytoid dendritic cells is essential for CpG-induced immune activation. A role for interleukin-2 transpresentation in dendritic cell-mediated T cell activation in humans, as revealed by daclizumab therapy. The structure of the interleukin-15 alpha receptor and its implications for ligand binding. A fundamental role for interleukin-21 in the generation of T follicular helper cells. Signals mediated by transforming growth factor-beta initiate autoimmune encephalomyelitis, but chronic inflammation is needed to sustain disease. Interleukin-21: a modulator of lymphoid proliferation, apoptosis and differentiation. Interleukin-2 receptor gamma chain mutation results in X-linked severe combined immunodeficiency in humans. Long-term chimerism and B-cell function after bone marrow transplantation in patients with severe combined immunodeficiency with B cells: a single-center study of 22 patients. Severe combined immunodeficiency due to a specific defect in the production of interleukin-2. Primary combined immunodeficiency resulting from defective transcription of multiple T-cell lymphokine genes. Development and function of T cells in mice rendered interleukin-2 deficient by gene targeting. Development and proliferation of lymphocytes in mice deficient for both interleukins-2 and -4. Cytoplasmic domains of the interleukin-2 receptor beta and gamma chains mediate the signal for T-cell proliferation. Deregulated T cell activation and autoimmunity in mice lacking interleukin-2 receptor beta. The common cytokine receptor gamma chain plays an essential role in regulating lymphoid homeostasis. Function and signal transduction mediated by the interleukin 3 receptor system in hematopoiesis. Cloning of an interleukin-3 receptor gene: a member of a distinct receptor gene family. Expression cloning of a receptor for human granulocyte-macrophage colony-stimulating factor. Cloning of the low-affinity murine granulocyte-macrophage colony-stimulating factor receptor and reconstitution of a high-affinity receptor complex. Cloning and expression of a gene encoding an interleukin 3 receptor-like protein: identification of another member of the cytokine receptor gene family. Characterization of critical residues in the cytoplasmic domain of the human interleukin-5 receptor alpha chain required for growth signal transduction. Hematopoiesis in mice lacking the entire granulocyte-macrophage colony-stimulating factor/interleukin-3/interleukin-5 functions. Functional inhibition of hematopoietic and neurotrophic cytokines by blocking the interleukin 6 signal transducer gp130. Biological activities and binding to the leukemia inhibitory factor receptor/gp130 signaling complex. High affinity interleukin-6 receptor is a hexameric complex consisting of two molecules each of interleukin-6, interleukin-6 receptor, and gp-130. Interleukin-11: a multifunctional growth factor derived from the hematopoietic microenvironment. Preclinical biology of interleukin 11: a multifunctional hematopoietic cytokine with potent thrombopoietic activity. Molecular cloning of two isoforms of a receptor for the human hematopoietic cytokine interleukin-11. The cholinergic neuronal differentiation factor from heart cells is identical to leukemia inhibitory factor. Leukemia inhibitory factor inhibits T helper 17 cell differentiation and confers treatment effects of neural progenitor cell therapy in autoimmune disease. Molecular cloning, expression and regional distribution of rat ciliary neurotrophic factor. Oncostatin M: a growth regulator produced by differentiated histiocytic lymphoma cells.
Myrtle Flag (Calamus). Doxepin.
- Dosing considerations for Calamus.
- What is Calamus?
- Are there safety concerns?
- Ulcers, gas, upset stomach, appetite stimulation, arthritis, strokes, and skin disorders.
- Are there any interactions with medications?
- How does Calamus work?
Source: http://www.rxlist.com/script/main/art.asp?articlekey=96757
Doxepin 25mg with visa
The primordial lymphoid progenitor has the potential to differentiate into B- or T-lymphocytes or natural killer cells anxiety symptoms skin rash generic 25mg doxepin with mastercard. A simplified scheme of B-cell development is presented as a background for discussion of Ig gene recombination. A graphic depicting the Ig-related proteins displayed on the surface at each stage is presented; at the bottom, the stage-dependent expression of recombination activating genes and terminal deoxynucleotidyl transferase-both important in V(D)J recombination-is schematically depicted. When immature B cells eventually also express surface IgD, they become "mature B cells" and migrate into the periphery, ready to be triggered by antigen exposure. Allelic Exclusion and Regulated V(D)J Recombination the previous description of B-cell development serves as a background to understand an explanation of allelic exclusion that was first proposed by Alt and colleagues110 and has been supported by subsequent experiments. According to this model the functional rearrangement of an L (or H) chain gene in a particular B cell would inhibit further L (or H) chain gene rearrangement in the same cell. If the inhibition occurred promptly after the first functional rearrangement, then two functional Igs could never be produced in the same cell. An initial nonproductive rearrangement would have no inhibitory effect, so recombination could continue until a functional product resulted or until the cell used up all its germline precursors. This effect was originally deduced from the rarity of -expressing cells without H chain gene rearrangement, suggesting that H chain expression is required for recombination. These are designated "sterile" transcripts because they cannot encode a protein, but they are required for V-J recombination. When this recombination ensues, the possibilities for functional and nonproductive V-J rearrangements resemble those discussed previously for the H chain. Thus in normal and malignant human B-lymphoid cells, -expressing cells generally have their genes in germline configuration, while in -expressing cells, genes are either rearranged (rarely) or deleted (most commonly) by recombination signal recombination events discussed previously in this chapter. Plausible models include either the selective suppression of recombination until all options on the Ig locus are exhausted or differences in the timing of the developmental programs controlling and accessibility. A widely accepted explanation for this locus specificity is provided by the "accessibility" model. One clue suggesting this model was that susceptibility to recombination and transcription of germline gene elements seem to be tightly correlated. We discuss a few important examples for each mechanism and refer to comprehensive review articles for an in-depth discussion. Surprisingly, the elements and mechanism for regulating expression during B- and T-cell development are distinct. Subnuclear Localization In general, inactive genes tend to be located in the periphery of nuclei, while active genes are recruited to a more central nuclear location. The IgH and Ig loci are located at the nuclear periphery in hematopoietic progenitors and pro-T cells, but move to central areas of the nucleus in pro-B cells. As mentioned previously, transcription of individual elements within Ig gene loci correlates well with their availability for V(D)J recombination at that stage. Distinct marks correlate well with actively transcribed and inactive (or repressed) gene loci. The developmental maturation from pro-B to pre-B cells is associated with progression from a locus that is largely methylated, nontranscribed, and nonrearranging to one that is hypomethylated, transcribed, and rearranging. Such distant segments are apparently brought close together in the nucleus, a process of "locus compaction" that loops out large cis Mediators of Accessibility and Looping All previously described properties are dependent on cisregulatory elements within the Ig loci, including classic promoters and enhancers. Individual promoters are present upstream of all V elements in all Ig loci, while the downstream D and J elements share a smaller number of promoters. Enhancers are present in each Ig locus, and they are thought to confer the transcriptional activation of each locus at the appropriate stage of B-cell development. Murine and IgH loci have intronic enhancers in the intron between J and C (iE and E, respectively), and all three loci have enhancers downstream of C coding regions. Transcriptional activation and the correlated locus "opening" is mediated by the recruitment of transcription factors that in turn recruit histone modifying and chromatin-remodeling enzymes. An enhancer may activate transcription of several genes within a given gene locus, but its effects may be deleterious if it can activate other nearby genes requiring different patterns of expression. To prevent enhancer function beyond appropriate domains, boundary elements known as insulators establish borders between gene loci that are differentially regulated. One such insulator apparently lies downstream of a complex of enhancers at the 3 end of the IgH locus, the 3-regulatory region, where it may protect genes further downstream from being regulated by the Ig enhancer elements. Most replacement of productively rearranged L or H chain genes likely serves to extinguish an antibody that was autoreactive, thus complementing two other mechanisms to silence autoantibodies: anergization and cell deletion by apoptosis. Early studies with transgenic autoantibodies suggested that anergy or deletion were the main fates of self-reactive B cells, but these conclusions may have depended on the nonphysiologic inability of the cells to silence the transgenic autoantibody by receptor editing. The recombination breakpoints in isotype switch recombination fall within repetitive "switch" (S) regions. Polymerase chain reaction amplification across either composite junction can be used to study switch recombination. The same defects are seen in patients with a homozygous defect in the human Aicda gene, a condition known as the hyper-IgM syndrome-2. The human Apobec1 and Aicda genes are genetically linked, both lying at chromosome 12p13. Other functional features of the protein include a dimerization domain, several phosphorylatable residues, segments affecting nuclear localization, and target sequences for association with other proteins. This reaction in germ cells may contribute to the most frequent germline point mutation observed in mammals: the transition from CpG to TpG. Perhaps to protect against such effects, miR155 is itself upregulated by the same signals that induce Aicda transcription in B cells. Movement between cytoplasm and nucleus is controlled by at least three independent mechanisms: active nuclear import mediated by an N-terminal nuclear localization signal,190 active nuclear export mediated by a C-terminal nuclear export signal,189,191 and a cytoplasmic retention mechanism apparent when active import and export mechanisms are both blocked. They include an upstream enhancer responsive to T-cell signals, a promoter, a regulatory region just downstream of exon 1, and an additional enhancer downstream of the Aicda gene. Persuasive evidence for somatic mutation was reported in the 1970s: analyses of V1 amino acid sequences of murine myeloma antibodies showed many instances of a particular prototype sequence, plus several variants containing unique single amino acid substitutions that could be explained by single nucleotide changes. The prototype was interpreted as reflecting the germline sequence, with the variants arising by somatic mutation. Many of these mutations reduce the affinity of the encoded antibody for antigen, 201 and some may induce autoantibody specificities. As clearance of the antigen lowers the antigen concentrations, only the cells displaying high affinity for antigen will be stimulated effectively; cells displaying lower-affinity antibodies or antibodies with affinity for self-antigens are subjected to programmed cell death ("apoptosis"). Some high-affinity cells become memory cells, persisting long after the initial antigen exposure, ready to respond to a subsequent antigen exposure with rapid production of high-affinity antibody. In this model, the driving force for affinity maturation- analogous to natural selection in the evolution of species-is selection for high-antibody affinity in the face of falling antigen concentration. Selection for high-affinity antibodies occurs because cells expressing high-affinity antibodies are most efficiently rescued from apoptosis. Surviving centrocytes may return to the dark zone to undergo several successive cycles of mutation and proliferation followed by selection. This model is supported by direct observation (by two-photon microscopy) of B cells moving between light zone and dark zone. Studies of the responses to p-azophenylarsonate (Ars), phosphorylcholine, influenza hemagglutinin, oxazalone, and several other antigens have all indicated that the initial response after primary immunization is established by antibodies showing no somatic mutation. About 1 week after immunization, mutated sequences begin to be observed, increasing during the next week or so. Many hybridomas made late in the immune response produce mutated antibodies with a higher antigen affinity than the unmutated (sometimes loosely called "germline") antibodies made early after immunization. The shift to higher affinity is a phenomenon long recognized at the level of (polyclonal) antisera and has been termed "affinity maturation. Error-prone repair inserts mispaired bases, which may become fi xed on one strand by replication, creating mutations (designated phase 2) from both A:T and G:C bps at some distance from the position of the original U:G mismatch. Polymerase characteristically inserts mismatched bases opposite T nucleotides, and individuals lacking this polymerase show the predicted abnormality: decreased mutations at A:T base pairs. This question is part of the larger issue of mutational targeting, as discussed below.
Syndromes
- Decreased alertness or orientation
- Have had a barium contrast media x-ray in the last 4 days
- Eczema (atopic dermatitis)
- Slowed respirations
- Asperger syndrome (like autism, but with normal language development)
- Kidney ultrasound
- Severe change in acid level of blood (pH balance), which leads to damage in all of the body organs
Cheap 10 mg doxepin
Identification of Igsigma and Iglambda in channel catfish anxiety symptoms and treatments discount 10mg doxepin fast delivery, Ictalurus punctatus, and Iglambda in Atlantic cod, Gadus morhua. Immunoglobulin light chain (IgL) genes in zebrafish: Genomic configurations and inversional rearrangements between (V(L)-J(L)-C(L)) gene clusters. Noncovalent association of heavy and light chains in Rana catesbeiana immunoglobulins. Immunoglobulin light chain class multiplicity and alternative organizational forms in early vertebrate phylogeny. Secretory antibody formation: conserved binding interactions between J chain and polymeric Ig receptor from humans and amphibians. The joining (J) chain is present in invertebrates that do not express immunoglobulins. Alpha, beta, gamma, and delta T cell antigen receptor genes arose early in vertebrate phylogeny. Identification of the pre-Tcell receptor alpha chain in nonmammalian vertebrates challenges the structure-function of the molecule. Multiple rearrangements in T cell receptor alpha chain genes maximize the production of useful thymocytes. Germlineencoded amino acids in the alphabeta T-cell receptor control thymic selection. Identification and characterization of T-cell antigen receptor-related genes in phylogenetically diverse vertebrate species. Origin and development of the gamma delta T-cell system in sheep: a critical role for the thymus in the generation of TcR diversity and tissue tropism. Diversity and evolution of T-cell receptor variable region genes in mammals and birds. Characterization of arrangement and expression of the T cell receptor gamma locus in the sandbar shark. Characterization of the immunoglobulin repertoire of the spiny dogfish (Squalus acanthias). Expression of individual immunoglobulin genes occurs in an unusual system consisting of multiple independent loci. Diverse immunoglobulin light chain organizations in fish retain potential to revise B cell receptor specificities. Hagfish humoral defense protein exhibits structural and functional homology with mammalian complement components. Lamprey lymphocyte-like cells express homologs of genes involved in immunologically relevant activities of mammalian lymphocytes. Somatic diversification of variable lymphocyte receptors in the agnathan sea lamprey. Antigen-receptor genes of the agnathan lamprey are assembled by a process involving copy choice. Structure of a lamprey variable lymphocyte receptor in complex with a protein antigen. A structural basis for antigen recognition by the T cell-like lymphocytes of sea lamprey. Lack of heterogeneity in antihapten antibodies of a phylogenetically primitive shark. Somatic variation precedes extensive diversification of germline sequences and combinatorial joining in the evolution of immunoglobulin heavy chain diversity. Hypermutation in shark immunoglobulin light chain genes results in contiguous substitutions. Somatic hypermutation and junctional diversification at Ig heavy chain loci in the nurse shark. Age, microbiota, and T cells shape diverse individual IgA repertoires in the intestine. First molecular and biochemical analysis of in vivo affinity maturation in an ectothermic vertebrate. The strength of B cell interaction with antigen determines the degree of IgM polymerization. Rhabdovirus infection induces public and private T cell responses in teleost fish. Transduction of binding affinity by B lymphocytes: a new dimension in immunological regulation. Comparative analyses of B cell populations in trout kidney and mouse bone marrow: establishing "B cell signatures. The nucleotide targets of somatic mutation and the role of selection in immunoglobulin heavy chains of a teleost fish. Activation-induced cytidine deaminase structure and functions: a species comparative view. Targets of somatic hypermutation within immunoglobulin light chain genes in zebrafish. Noninbred axolotls used the same unique heavy chain and a limited number of light chains for their anti-2,4-dinitrophenyl antibody responses. Activation by mitogens and superantigens of axolotl lymphocytes: functional characterization and ontogenic study. What limits affinity maturation of antibodies in Xenopus-the rate of somatic mutation or the ability to select mutants Localization and differential expression of activation-induced cytidine deaminase in the amphibian Xenopus upon antigen stimulation and during early development. In vivo and in vitro techniques for comparative study of antiviral T-cell responses in the amphibian Xenopus. Hapten-binding capacity and population heterogeneity of chicken anti-Dnp IgG antibody. Evolution of genetic networks underlying the emergence of thymopoiesis in vertebrates. The development of primary and secondary lymphoid tissues in the nurse shark Ginglymostoma cirratum: B-cell zones precede dendritic cell immigration and T-cell zone formation during ontogeny of the spleen. Complex expression patterns of lymphocyte-specific genes during the development of cartilaginous fish implicate unique lymphoid tissues in generating an immune repertoire. Salmonid T cells assemble in the thymus, spleen and in novel interbranchial lymphoid tissue. Characterization of rainbow trout terminal deoxynucleotidyl transferase structure and expression. Defining terminally differentiating B cell populations in rainbow trout immune tissues using the transcription factor XbpI. B lymphocytes from early vertebrates have potent phagocytic and microbicidal abilities. Ultrastructural changes in the spleen of the natterjack, Bufo calamita, after antigenic stimulation. Response to skin allografts in Xenopus laevis following thymectomy at early stages of lymphoid organ maturation. The ontogeny of diversification at the immunoglobulin heavy chain locus in Xenopus. Isolation and characterization of the Xenopus terminal deoxynucleotidyl transferase. Antibodies, immunoglobulin genes and the bursa of Fabricius in chicken B cell development. A hyperconversion mechanism generates the chicken light chain preimmune repertoire. Formation of the chicken B-cell repertoire: ontogenesis, regulation of Ig gene rearrangement, and diversification by gene conversion. Efficient antibody diversification by gene conversion in vivo in the absence of selection for V(D)J-encoded determinants. Positive selection of the peripheral B cell repertoire in gut-associated lymphoid tissues. Evolutionarily conserved and divergent regions of the autoimmune regulator (Aire) gene: a comparative analysis. Comparative genomic analysis of the proteasome beta5t subunit gene: implications for the origin and evolution of thymoproteasomes. Physical and genetic mapping of the rainbow trout major histocompatibility regions: evidence for duplication of the class I region.
Purchase doxepin 75mg mastercard
Innate immunity defines the capacity of antiviral T cells to limit persistent infection anxiety vs heart attack cheap doxepin 25mg otc. Transforming growth factorbeta controls T helper type 1 cell development through regulation of natural killer cell interferon-gamma. Cancer immunosurveillance and immunoediting: the roles of immunity in suppressing tumor development and shaping tumor immunogenicity. Critical role for tumor necrosis factor-related apoptosis-inducing ligand in immune surveillance against tumor development. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: an 11-year follow-up study of a general population. Interleukin-2 enhances the depressed natural killer and cytomegalovirus-specific cytotoxic activities of lymphocytes from patients with the acquired immune deficiency syndrome. Inter-relationship among macrophages, natural killer cells and neutrophils in early stages of Listeria resistance. Natural killer cell depletion enhances virus synthesis and virus-induced hepatitis in vivo. Pathogenesis of murine cytomegalovirus infection in natural killer cell-depleted mice. Requirement for natural killer cell-produced interferon gamma in defense against murine cytomegalovirus infection and enhancement of this defense pathway by interleukin 12 administration. Natural killer cells utilize both perforin and gamma interferon to regulate murine cytomegalovirus infection in the spleen and liver. Natural immunity: a T-cellindependent pathway of macrophage activation, defined in the scid mouse. Early gamma interferon production by natural killer cells is important in defense against murine listeriosis. Natural killer cells in antiviral defense: function and regulation by innate cytokines. Adoptive transfer studies demonstrating the antiviral effect of natural killer cells in vivo. Noncytolytic control of viral infections by the innate and adaptive immune response. Interleukin-15 as an activator of natural killer cell-mediated antiviral response. Innate immune response of the human host to exposure with herpes simplex virus type 1: in vitro control of the virus infection by enhanced natural killer activity via interleukin-15 induction. A protective role of interleukin-15 in a mouse model for systemic infection with herpes simplex virus. Control of natural killer cellmediated innate resistance against the intracellular pathogen Listeria monocytogenes by gamma/delta T lymphocytes. Alpha-galactosylceramide as a therapeutic agent for pulmonary Mycobacterium tuberculosis infection. Cellular expression and crystal structure of the murine cytomegalovirus major histocompatibility complex class I-like glycoprotein, m153. Killing of human immunodeficiency virusinfected primary T-cell blasts by autologous natural killer cells is dependent on the ability of the virus to alter the expression of major histocompatibility complex class I molecules. Cmv-1, a genetic locus that controls murine cytomegalovirus replication in the spleen. The effect of the Cmv-1 resistance gene, which is linked to the natural killer cell gene complex, is mediated by natural killer cells. Vital involvement of a natural killer cell activation receptor in resistance to viral infection. Susceptibility to mouse cytomegalovirus is associated with deletion of an activating natural killer cell receptor of the C-type lectin superfamily. Murine cytomegalovirus is regulated by a discrete subset of natural killer cells reactive with monoclonal antibody to Ly49h. Transgenic expression of the activating natural killer receptor Ly49H confers resistance to cytomegalovirus in genetically susceptible mice. Murine cytomegalovirus m157 mutation and variation leads to immune evasion of natural killer cells. Epistasis between mouse Klra and major histocompatibility complex class I loci is associated with a new mechanism of natural killer cell-mediated innate resistance to cytomegalovirus infection. Coordinate expression of cytokines and chemokines by natural killer cells during murine cytomegalovirus infection. Induction of bystander T cell proliferation by viruses and type I interferon in vivo. Unconventional repertoire profile is imprinted during acute chikungunya infection for natural killer cells polarization toward cytotoxicity. Natural killer cells and T cells induce different types of skin reactions during recall responses to haptens. Dendritic cells in pathogen recognition and induction of immune responses: a functional genomics approach. Dendritic cell maturation by innate lymphocytes: coordinated stimulation of innate and adaptive immunity. Natural killer cell behavior in lymph nodes revealed by static and real-time imaging. Granulated metrial gland cells: hypotheses concerning possible functions during murine gestation. Intrinsic susceptibility of mouse trophoblasts to natural killer cell-mediated attack in vivo. Interferon gamma contributes to initiation of uterine vascular modification, decidual integrity, and uterine natural killer cell maturation during normal murine pregnancy. Expression of angiogenic growth factors by uterine natural killer cells during early pregnancy. Hierarchy of resistance to cervical neoplasia mediated by combinations of killer immunoglobulin-like receptor and human leukocyte antigen loci. Natural killer cell dysfunction in patients with systemic-onset juvenile rheumatoid arthritis and macrophage activation syndrome. Bortezomib treatment to potentiate the anti-tumor immunity of ex-vivo expanded adoptively infused autologous natural killer cells. Primary nasal natural killer cell lymphoma: long-term treatment outcome and relationship with the International Prognostic Index. A semi-invariant Valpha10+ T cell antigen receptor defines a population of natural killer T cells with distinct glycolipid antigen-recognition properties. Structure-activity relationship of alpha-galactosylceramides against B16-bearing mice. A modified alpha-galactosyl ceramide for staining and stimulating natural killer T cells. Structural analysis of two glycosphingolipids from the lipopolysaccharide-lacking bacterium Sphingomonas capsulata. Natural killer T cells recognize diacylglycerol antigens from pathogenic bacteria. Invariant natural killer T cells recognize glycolipids from pathogenic Gram-positive bacteria. Mechanisms imposing the Vbeta bias of Valpha14 natural killer T cells and consequences for microbial glycolipid recognition. Lysosomal alpha-galactosidase controls the generation of self lipid antigens for natural killer T cells. Sensitive detection of isoglobo and globo series tetraglycosylceramides in human thymus by ion trap mass spectrometry. Sensitive quantitation of isoglobotriaosylceramide in the presence of isobaric components using electrospray ionization-ion trap mass spectrometry. Normal development and function of invariant natural killer T cells in mice with isoglobotrihexosylceramide (iGb3) deficiency. Structure and function of a potent agonist for the semi-invariant natural killer T cell receptor. Recognition of beta-linked self glycolipids mediated by natural killer T cell antigen receptors. Intrathymic proliferation wave essential for Valpha14+ natural killer T cell development depends on c-Myc.
Generic 75mg doxepin overnight delivery
Transmission-blocking immunity has been induced in vivo by immunization with gametes of avian anxiety exercises discount doxepin 10mg with visa, rodent, and monkey malarias. Several potential transmission-blocking vaccine candidates have been identified, and the genes encoding these surface proteins have been isolated and sequenced, but their production as recombinant proteins is hampered by failure to recreate the highly complex tertiary structures that are the targets of inhibitory antibodies. Furthermore, for those antigens expressed only by invertebrate stages of the parasite, lack of natural boosting may prevent maintenance of sufficiently high antibody titers. Clearly, an optimal vaccine against malaria would need to target multiple antigens and induce immunity against all stages. However, by targeting certain antigens confined to asexual blood stages, the induction of an adult-like immune status among high-risk infants in sub-Saharan Africa could greatly diminish severe disease and death caused by P. T-Cell Vaccines Vaccination Against Leishmaniasis Vaccines against intracellular parasites will need to induce long-lived cellular immune responses. An inherent problem with most nonliving vaccines is their relative inefficiency in generating and/or sustaining these sorts of cellular responses. A major advance in T-cell vaccine development was the demonstration that proteins derived from L. The gold standard of the protection that can be achieved by vaccination in mice, and the only vaccination strategy against leishmaniasis that has worked so far in humans is " leishmanization," which is based on the lifelong convalescent immunity that is acquired following induction of a lesion at a selected site with a cutaneous strain of L. The nature of the acquired resistance that develops following healing of a primary lesion seems especially important to consider as only healed mice were protected against L. Although the efficacy of live attenuated vaccines against needle challenge in mice has been shown,431 none have been evaluated using infected sand flies and more generally pose greater difficulties in standardization and delivery in field settings. Whereas a number of experimental vaccines were shown to reduce the tissue inflammation and parasitism associated with late chronic phase infections, it does not seem that infection itself, or even infection and drug cure, confers sterilizing immunity against reinfection,146 and there is so far no evidence that a vaccine can achieve a better result. Furthermore, the immune response seems to be focused on epitopes encoded by genes of the large and strain variant trans-sialidase gene family,436 which would require that a massive number of target epitopes be included in an effective vaccine. Thus, the rationale for the development of a safe and effective vaccine against human Chagas disease remains suspect, particularly as vector control methods have proven to be so highly effective. Vaccines to prevent toxoplasma infection are needed primarily to protect livestock and prevent transmission to humans from felines. These findings offer new promise to peptide-based approaches for vaccination against T. A recombinant polypeptide representing the central repeat and C-terminal portions of the P. The preliminary results of this trial confi rm that the vaccine has approximately 50% efficacy against clinical malaria attacks and against malaria-related hospital admissions,443 but the duration of protection is still an area of concern. Although protection in human trials has been less convincing, delays in time to emergence of blood-stage parasites have been consistently obtained, suggesting that the vaccination substantially reduces, but does not eliminate, liver stage parasites. Vaccines Against Helminths Infection with helminthic parasites remains a significant health problem in many tropical countries. Whereas control measures are available in some areas, in most cases, patients living in endemic regions are quickly reinfected. Therefore, vaccines that reduce parasite and/or egg burdens would be a valuable tool to complement existing disease prevention programs and could represent a less costly and more practical approach than repeated chemotherapy. Although many subunit vaccines have been described and tested in various animal models, suboptimal levels of protection have hindered the development of all but a few of these candidate vaccines. However, in order to effectively harness and implement these advances, it will be necessary to fully understand the mechanisms of resistance to helminth parasites. Vaccine models using radiation-attenuated larval parasites have provided the best examples of successful immunization against helminths. Other antigens of interest include paramyosin (Sm97), IrV5 (myosin-like 62kDa protein), triose phosphate isomerase, Sm23, the integral membrane protein tetraspanin-2, and Sm14, a fatty acidbinding protein. Because these pathogens are so well adapted to their hosts, it may be easier and more efficient to design immune interventions that prevent parasite-induced immunopathology rather than eliminate the infection itself. Although this approach will not lead to eradication of the parasite, it would likely reduce or alleviate the health consequences of infection. The feasibility of antipathology vaccines was demonstrated in a murine models of schistosomiasis. Importantly, several immunodominant egg antigens have been described; 449,450 thus, it may be possible to design recombinant antipathology vaccines that duplicate the promising results produced with crude parasite extracts. It is hoped, however, that ongoing improvements in vaccination technology, combined with greater knowledge of the mechanisms controlling resistance, will allow development of more efficacious and better defined vaccines for these complex organisms. Indeed, although once regarded as a less sophisticated topic than protozoan immunology, the study of immunity to helminths has undergone a major resurgence in part because of the growing appreciation of the phylogenetic uniqueness of worm pathogens and their host interaction. Research on the immune response to helminths has also been stimulated by the growing interest in neglected tropical diseases, where worm infections account for the most affected individuals. Finally, there has been a growing awareness of the important evolutionary role played by helminth infection in conditioning the mammalian immune system, in the maintenance of immunologic homeostasis at mucosal tissue barriers, and of the likely impact of the loss of this symbiotic relationship on immune function due to modern improvements in human hygiene. Interestingly, many of these projects98,438,439 have brought together molecular and immunoparasitologists with the common goal of simultaneously identifying both parasite epitopes/virulence determinants and their host receptors and signaling pathway targets. The findings gained from such studies should provide important groundwork for future systems biology approaches that seek to develop a broader vision of the interaction of parasites with the immune system. Another important development since the publication of the last version of this chapter is the increased emphasis on the study of the human immune response to parasitic infection. The most notable advances have occurred in the field of malaria, which benefits from a wealth of clinical material and the ability to use peripheral blood as a physiologically relevant source of immune cells. The study of human malaria is yielding novel insights into the nature of B-cell effector and memory responses to the parasite as well as the role of innate cellular function in regulating disease outcome. Nevertheless, having a defined antigen vaccine in which at least a proportion of the immunized individuals become protected offers a rare opportunity to identify correlates of immunity to malaria triggered by a single immunogen through the use of a "systems vaccinology" approach. It is hoped that exciting new approaches and opportunities of this kind will bring us closer to our ultimate goal of protecting human populations against the scourge of parasitic disease. Wynn, David Sacks, and Alan Sher are all supported by the intramural research program, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Bethesda, Maryland. We thank Tom Nutman, Dragana Jankovic, and Thirumalai Ramalingam for helpful discussions. Although viruses are minute particles composed only of a genome surrounded by a few proteins, they have a fiendish way of wreaking havoc not only on humans, as everyone who ever had the flu knows, but also on animals, plants, and even bacteria. Each year, more people die of viral infections than of natural disasters such as hurricanes, earthquakes, and tsunamis combined, or even manmade tragedies such as war. For example, it is estimated that variola major, the causative agent for smallpox, killed nearly half of the population of Native Americans after the virus was introduced into the western hemisphere by European colonizers. The Spanish Flu caused by an H1N1 influenza A virus caused the death of 50 to 100 million humans between 1918 and 1919, which is well in excess of the 16 million casualties of World War I. Not only do viruses cause acute or chronic infections with potentially fatal outcome, but they also contribute to other diseases. Viruses are associated with 20% of cancers, they have been implicated in the pathogenesis of human arteriosclerosis and autoimmune diseases, and they are linked to an overall reduction in life expectancy. Viruses, which range in size from 10 to 300 nm in diameter with genomes of minimally 2 kilobases to over 1. Once a cell becomes infected, viruses hijack its transcription and translation machinery to promote their own replication. The physiologic functions of the infected cell are disrupted as it is being turned into a virus production facility. This fight initially takes place intrinsically within the infected cells but then rapidly spreads extrinsically once the immune system has been alerted. In turn, many viruses mount defenses against the attack from their host by encoding proteins that actively subvert innate and adaptive immune responses. In acute virus infections, the fight between virus and host literally lasts until the death of one of the adversaries. In chronic infections, a truce is eventually reached where virus and host coexist, generally at the expense of the well-being of the latter. Over the last 50 years, on average two new species of human viruses have been discovered annually; one can expect that this number will continue to rise. We are not certain about the origin of viruses, although we know from ancient texts as well as more modern data-driven genomic analyses that viruses have been around for a very long time. The constant discovery of new viruses may simply reflect improvements of detection technologies that traditionally were based on cell culture and that are now being replaced with high-throughput genomics.
