

Cheap minipress online amex
Long-term behavioral impairment following acute embryonic ethanol exposure in zebrafish antiviral influenza drugs generic minipress 1mg overnight delivery. Live imaging of adult neural stem cell behavior in the intact and injured zebrafish brain. Dystrophin is required for the formation of stable muscle attachments in the zebrafish embryo. Use of alternative assays to identify and prioritize potential developmental and neurotoxicity of emerging organophosphorous flame retardants. The role of the aryl hydrocarbon receptor pathway in mediating synergistic developmental toxicity of polycyclic aromatic hydrocarbons to zebrafish. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Ethanol effects on the developing zebrafish: neurobehavior and skeletal morphogenesis. Translating rodent behavioral repertoire to zebrafish (Danio rerio): relevance for stress research. Axonal transport defects in a mitofusin 2 loss of function model of Charcot-Marie-Tooth disease in zebrafish. Two Cyp19 (P450 aromatase) genes on duplicated zebrafish chromosomes are expressed in ovary or brain. Accelerated optical projection tomography applied to in vivo imaging of zebrafish. Regeneration, plasticity, and induced molecular programs in adult zebrafish brain. Neurobehavioral impairments caused by developmental imidacloprid exposure in zebrafish. Analysis of protein expression in developmental toxicity induced by MeHg in zebrafish. Identification of a conserved region requried fro hormone dependent transcriptional actiation by steroid-hormone receptors. Thyroid hormone receptors in two model species for vertebrate embryonic development: chicken and zebrafish. Spatial regionalization and heterochrony in the formation of adult pallial neural stem cells. Developmental exposure to organophosphate flame retardants elicits overt toxicity and alters behavior in early life stage zebrafish (Danio rerio). Heritable targeted gene disruption in zebrafish using designed zinc-finger nucleases. Temproal colinearity and the phylotypic progressionda basis for the stability of a vertebrate bauplan and the evoluiton of morphologies through heterochrony. Zebrafish provide a sensitive model of persisting neurobehavioral effects of developmental chlorpyrifos exposure: comparison with nicotine and pilocarpine effects and relationship to dopamine deficits. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. A behavioral assay to measure responsiveness of zebrafish to changes in light intensities. Embryonic alcohol exposure impairs associative learning performance in adult zebrafish. Some principles of organization of spinal neurons underlying locomotion in zebrafish and their implications. Comparative analysis of goitrogenic effects of phenylthiourea and methimazole in zebrafish embryos. Identification and characterization of androgen-responsive genes in zebrafish embryos. Phenotype-driven chemical screening in zebrafish for compounds that inhibit collective cell migration identifies multiple pathways potentially involved in metastatic invasion. Use of a high-throughput screening approach coupled with in vivo zebrafish embryo screening to develop hazard ranking for engineered nanomaterials. Drinks like a fish: zebra fish (Danio rerio) as a behavior genetic model to study alcohol effects. Identification and developmental expression of the full complement of Cytochrome P450 genes in Zebrafish. Insertional mutagenesis in zebrafish rapidly identifies genes essential for early vertebrate development. Transgenic zebrafish reveal tissue-specific differences in estrogen signaling in response to environmental water samples. Genes controlling and mediating locomotion behavior of the zebrafish embryo and larva. Linking genes to brain, behavior and neurological diseases: what can we learn from zebrafish Whole-organism screening for gluconeogenesis identifies activators of fasting metabolism. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. The transcriptional response to oxidative stress during vertebrate development: effects of tert-butylhydroquinone and 2,3,7,8-tetrachlorodibenzo-p-dioxin. Comparative embryotoxicity and proteotoxicity of three carrier solvents to zebrafish (Danio rerio) embryos. Adenomatous polyposis coli-deficient zebrafish are susceptible to digestive tract neoplasia. Systematic evaluation of nanomaterial toxicity: utility of standardized materials and rapid assays. Sublethal exposure to crude oil during embryonic development alters cardiac morphology and reduces aerobic capacity in adult fish. Brain and gonadal aromatase as potential targets of endocrine disrupting chemicals in a model species, the zebrafish (Danio rerio). Reversed scototaxis during withdrawal after daily-moderate, but not weekly-binge, administration of ethanol in zebrafish. Interactions between chemical and climate stressors: a role for mechanistic toxicology in assessing climate change risks. Defects in cardiac function precede morphological abnormalities in fish embryos exposed to polycyclic aromatic hydrocarbons. Cardiac toxicity of 5-ring polycyclic aromatic hydrocarbons is differentially dependent on the aryl hydrocarbon receptor 2 isoform during zebrafish development. Comparative transcriptome analysis reveals vertebrate phylotypic period during organogenesis. Acute and developmental behavioral effects of flame retardants and related chemicals in zebrafish. Identification of a neural stem cell in the adult mammalian central nervous system. Using zebrafish to uncover the genetic and neural basis of aggression, a frequent comorbid symptom of psychiatric disorders. A transposon-mediated gene trap approach identifies developmentally regulated genes in zebrafish. Reconstruction of zebrafish early embryonic development by scanned light sheet microscopy. Caffeine neuroprotects against dexamethasone-induced anxiety-like behaviour in the zebrafish (Danio rerio).
Purchase generic minipress pills
Clinical correlates associated with hypothalamic lesions encompass disorders of water imbalance anti viral fungal fighter order 2 mg minipress, body temperature regulation, caloric balance, alertness, sleep, memory, and emotional behavior. It contains the lower motor neurons for the muscles of the head and receives sensory input from this region. It also serves as a pathway for most of the ascending and descending fiber tracts between the spinal cord and the brain. The midbrain segment of the brain stem contains nuclei important for generalized motor control. A fourth major activity of the brain stem involves integrative activity that controls respiration, cardiovascular activity, and regulation of the level of consciousness. Each region is anatomically divided into a rostral (front) and caudal (rear) segment. The reticular formation is a diffusely organized series of nuclei and tracts that forms the central core of the three brain stem regions. The reticular formation is involved in motor, sensory, and visceral control of processes such as respiration and cardiovascular activity and control of consciousness. The medial lemniscus ascends through all regions of the brain stem and contains fibers from the contralateral neurons in the dorsal spinal cord, which receive sensory inputs from the periphery. The mass of neurons forming the third cranial nerve (occulomotor) lies in the midportion of the rostral midbrain. The fourth cranial nerve (trochlear) is located in the midregion of the caudal midbrain. The superior cerebellar peduncle is the major outflow from the cerebellum to the thalamus and red nucleus. Fibers from the superior cerebellar peduncle surround the midline and begin to cross over or decussate at this level. Complete immobility except for eye movement (akinetic mutism) and decerebrate rigidity may also occur with damage to the midbrain. The pons is made up of a central core, the reticular formation, containing the serotonergic raphe nuclei, which allow communication between the cerebellum and the cerebral cortex, and the noradrenergic locus ceruleus, which projects to virtually all areas of the cerebral cortex and spinal cord. The pons is attached to the cerebellum at the superior and middle cerebellar peduncles ("Cerebellum" section). The nucleus of the fifth cranial nerve (trigeminal) is located in the rostral pons. Like the midbrain and pons, the medulla contains a reticular core surrounded by fiber tracts from ascending and descending pathways, and communication pathways to and from the cerebellum. The caudal medulla is the site of the decussation (crossing) of fibers in the corticospinal tract (the decussation of the pyramids) and fibers in the medial lemniscus. The lateral reticular nucleus and inferior olivary nuclear complex in the medulla are important sources of input to the cerebellum via the inferior cerebellar peduncle. The lateral reticular nucleus receives input from the spinal cord, cranial nerves, and cerebral cortex, which it relays to the cerebellum. The neurons in the inferior olive are the source of the climbing fibers in the cerebellum. The medulla is the site for several important cranial nerves that innervate the pharynx, tongue, larynx, auditory canal, and nerve plexi in several major organ systems including the heart, lungs, and intestines. Medullary lesions can thus adversely affect these systems and the exact neuropathological phenotype will depend on the area of the medulla that is compromised. It functions to coordinate motor movement, maintain posture and balance, help execute fine motor skills such as writing, and aid in refining certain properties of movement such as trajectory, velocity, and acceleration. The granule cells of the cerebellum are particularly sensitive to damage by methyl mercury. The cerebellum is attached to the brain stem via three peduncles: superior, middle, and inferior. The superior cerebellar peduncle carries most of the outflow of the cerebellum, whereas the middle and inferior peduncles carry most of the input. The outflow of the cerebellum via the superior cerebellar peduncle projects to the red nucleus (midbrain), thalamus (diencephalon), the inferior olivary nucleus, and the reticular formation of the medulla. From these foci, information can be relayed to the motor cortex and spinal cord to facilitate motor adjustments that result in fine motor skills. The outflow of the cerebellum is, in turn, modified by the input to the cerebellum. Mossy and climbing fibers, and granule cells, are glutamatergic and, therefore, are excitatory. Information leaving the cerebellum occurs through the Purkinje cell that first synapses onto deep cerebellar nuclei. The spinal cord begins at its junction with the medulla and extends the length of the spinal column to the first lumbar vertebra. The cord is surrounded by a continuation of the three membranes or meninges that surround the brain: pia, arachnoid, and dura mater. There are two dorsal and two ventral enlargements of the gray matter known as the dorsal and ventral horns. The cord is divided into a right and left half by a dorsal and ventral median fissure. The spinal cord is divided into segments referred to as cervical, thoracic, lumbar, and sacral and corresponds with the attached spinal nerves ("Spinal nerves" section). Projections from the neurons in the dorsal horn of the cord give rise to tracts (spinothalamic, spinocerebellar, spino-olivary, spinotectal), which project to various parts of the brain, conveying sensory information from the periphery. Each dorsal horn is capped by a specialized area of neurons called the substantia gelatinosa of Rolando. The anterior or ventral horn contains the cells of origin (anterior motor neurons) of the ventral root. Afferent sensory input is received by neurons in the dorsal root ganglia, which send projections to cells in the posterior (P) or dorsal horn via the dorsal root (D). The ventral root (V) contains efferent projections from motor neurons in the anterior (A) or ventral horn and preganglionic fibers from neurons in the intermediate gray zone (I). Monosynaptic reflexes such as the knee jerk involve two neurons: a dorsal root or sensory neuron, which receives stimuli from the periphery and directly projects onto a motor neuron in the anterior horn. The axonal projection from the motor neuron ends in a neuromuscular spindle on the muscle. Polysynaptic reflexes involve spinal cord interneurons, which relay information from sensory neurons to motor neurons. Fundamentals of the Structure and Function of the Nervous System 19 skeletal muscles. Damage or death of anterior horn cells (lower motor neurons) leads to flaccid paralysis (limp, weak muscle tension, and inability to move) and eventually atrophy of the muscles innervated by these neurons. The lateral columns that are found in the intermediate area of gray matter contain preganglionic cells of the autonomic nervous system ("Autonomic nervous system" section) and also exit the cord via the ventral root. The cord is also the center for many monosynaptic reflex actions such as the knee jerk reflex seen in response to tapping the patellar tendon. For this reflex to occur, sensory information must reach the dorsal horn via the sensory afferents and dorsal root ganglia. Projections from the dorsal root ganglia synapse directly on motor neurons in the ventral horn, which in turn project to leg muscles. Most other polysynaptic reflexes involve spinal cord interneurons, which relay information to the motor neurons in the anterior horn. In addition to the numerous sensory tracts ascending from the spinal cord to the brain, the cord white matter contains the pathways descending from the brain (corticospinal, reticulospinal, etc. Fibers from these descending tracts synapse on motor neurons either directly or via interneurons. Disruption of these upper motor neuron pathways results in a spastic paralysis (increased muscle tension and inability to move). Distinction between flaccid and spastic paralysis can aid in locating the site of nerve cell damage. Many ascending tracts are then formed to convey sensory information to the brain ("Spinal cord" section). The ventral root of the spinal nerve contains the motor fibers of anterior motor neurons in the ventral horn, which pass to the skeletal muscles. In the thoracic and lumbar regions of the cord, the ventral root also contains preganglionic sympathetic fibers from the intermediate lateral cord to the autonomic sympathetic ganglia. In the sacral region, the ventral root carries preganglionic parasympathetic fibers from neurons in the intermediate gray of the cord to parasympathetic ganglia that innervate the pelvic region ("Autonomic nervous system" section). A summary of the 12 cranial nerves, their sites of origin, major destination(s), and functions is given in Table 3.
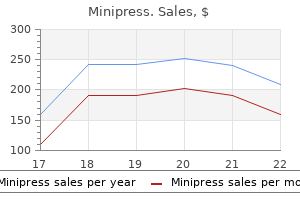
Order minipress overnight delivery
Relationship with the particular biological phenomenon is explained by establishing the relationship between interactional energies and biological activity hiv infection graph buy minipress uk. Also, its analysis should also use unambiguous machine learning method to generate the model. Applicability domain should also be well defined to have a better understanding of the predictive results. Rigorous methods should be implemented to estimate the robustness of the model, goodness of fit, and predictive accuracy. Mechanistic understanding of the results in turn increases the applicability of the results. The underlying interaction can be predicted using molecular modeling and docking techniques. For performing risk assessments, modeling and docking approaches can provide the approximations of toxicant and biological target interactions. This target interaction helps researchers in the identification of the mechanism of toxicity. A chemical toxicant (drug or environment chemical) has the potential to cause toxicity by various mechanisms, thus understanding of toxicant interaction with multiple targets is a differential step. The structure of macromolecular target can be either known or unknown, and structural conformation can be modeled using fragments conformations from previously known structures using homology modeling techniques (Shen et al. Toxicant structural information can be evaluated regarding the descriptors (Hong et al. This method was initially involved in the identification of new molecular entities for pharmaceutical uses (Coupez and Lewis, 2006) and predicted interactions were extensively studied for the understanding of the specific biological activity. However, the same methodology can be employed for the understanding of specific toxicity behavior and models for predicting toxicity can be developed (Rabinowitz et al. Predictive toxicity models using docking techniques have been widely used in drug discovery for efficacy. In toxicology, the technique can be used to investigate both on-target and off-target adverse events. A major limitation of this technique is that docking algorithm relays upon the availability of crystal structures of the target. However, ligand-based pharmacophore approaches or homology modeling-based approaches (Shen et al. This predictive modeling approach helps in the understanding of unknown biological activity underlying mode of 346 Computational Toxicology toxicity. With the use of docking methods, the likelihood of binding and probable mode of binding can be predicted for a particular macromolecular target for the library of toxicants (Jorgensen, 2004; Rabinowitz et al. Binding site can be determined by employing various algorithms and their location varies from protein to protein (Thakkar et al. Binding simulation algorithm takes account of ligand flexibility and binding site flexibility while predicting the mode and likelihood of binding. The output predicting the interaction of toxicant with macromolecular targets represented in terms of the binding energy. Docking methodology can also be used in identifying agonist and antagonist for understanding direct or indirect interactions as depicted in the study conducted by Ng et al. It also tests the ligands and decoy molecule to test the ability of identifying agonist and antagonist structures. It further tests individual model as well as combined models using enrichment factors. However, the challenge scientists face that they do not have enough data points to build predictive models. In this regard, grouping and read-across methods are beneficial to the assessment of toxicity profile. This approach is useful in filling of data gaps for predicting the safety concerns. Read-across method groups the substances with similar chemical structure, thus all chemicals in the group exhibit similar physicochemical profiles. Therefore, those groups of compounds will also have similar toxicokinetics/dynamics profile (Schultz et al. With the use of the read-across methods, researchers can extrapolate toxicity information from the one chemical (source) and fill the toxicity information of another chemical (target) with similar structure. Toxicity of many untested chemicals (target) can be assessed using the results from same toxicity endpoints from similarly tested compounds (source). Read-across based on the categories has been demonstrated to identify chemicals with hazardous property (Enoch and Roberts, 2013). One of the limitations of read-across is to identify comparatively less dangerous substances. Therefore, it is important that similarity parameters are carefully established to achieve better accuracy. The similarity parameters could be based on the structure and chemical classification, physiochemical properties, functional group and fragments, toxicokinetic and metabolic similarity, and potential of formation of metabolic products. Mechanistic information as well as in vivo and in vitro endpoints can also be incorporated to identify toxicant. There are several recognized approaches to reduce the uncertainty in the prediction. Risk can be reduced by increased number of analogs in source set, improving the data with greater quality of in vivo endpoints, including the points with severe in vivo hazard, including more biological causality evidence, and so on. There are several identified approaches to reduce the uncertainty in the prediction. Uncertainty can be reduced by increase number of analogues in source set, improving the data with greater quality of in vivo endpoints, including the points with severe in vivo hazard, including more biological causality evidence, and so on. Ultimately, read-across prediction can be relevant to make regulatory recommendations. Ongoing efforts in the field of developmental toxicity assessments try to identify and evaluate testing strategies for an understanding of the mechanism of toxicity. However, current testing methods have limited availability of information about in utero exposure to chemicals and drugs, the risks of adsorption in children, and the risks from lifelong exposure and the extent of exposure. Several computational toxicological-related resources discussed in this article are intended to provide diverse data types to empower an integrative approach for predictive modeling. It has also been demonstrated that integrative approach for predictive modeling is much more significant. Thus, these computational toxicological resources are precious for advancing the understanding of the field. Next-generation toxicity estimation techniques require a mechanism of toxicity information for estimating human risks. However, with the advances in high-throughput technologies, such as omics and next-generation sequencing, we can perform systems biology-level analysis to improve the prediction of the likelihood of an adverse outcome. Text mining techniques are beneficial in identifying the hidden relation between toxicant and adverse outcome from literature, and results can be enhanced with the extensive use of ontology. Computational methods are diverse and many of them have been adopted by computational toxicology such as data mining, chemoinformatics, and machine learning methods for understanding developmental toxicity. The article also listed examples demonstrating successful execution of several predictive computational toxicological-related projects and shed light on the new paradigm. Such data provides the prospect of replacing speculation about toxicology with confirmed facts. As a model can never be any better than the data it is fit to , experimental errors will be encoded into models. Likewise, errors can be made executing complex pipelines of algorithms used to extract and interpret experimental data. Discipline is necessary for adhering to well-known principles in model development that avoid error. This article provided the importance of mathematical logic in building valid models, then the enormous flexibility in methods available, model validation, and the increasingly important topic of model reliability and applicability domain. An immediately apparent risk is false discovery when, for example, tens of thousands of hypotheses are simultaneously tested. Thus, model validation is important where model validity is often tested by building models from a part of the data to predict a portion not used in training. Thus, these models are capable of performing both dimension reduction and data labeling according to its latent variables. With the use of latent modeling, approaches, large documents can not only be structured and rank ordered but also be useful for clustering analysis.
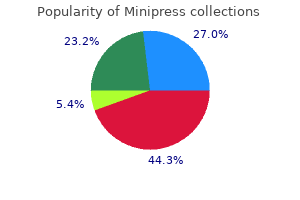
Order minipress amex
Reproductive lesions in female Harlan SpragueDawley rats following two-year oral treatment with dioxin and dioxin-like compounds hiv infection rate spain minipress 2 mg low price. Azole fungicides affect mammalian steroidogenesis by inhibiting sterol 14 alpha-demethylase and aromatase. Comparative studies on the increase of uterine weight and related mechanisms of cadmium and p-nonlyphenol. Juanmahel Davila, Quanx Li, and Indrani C Bagchi updated the author list, abbreviation list, keywords, introductory sections. The generation of new offspring requires gametes from both parents, followed by successful fertilization and embryogenesis. Establishment of a functional reproductive system is essential for the production of viable gametes. In females, a functional reproductive system consists of ovaries, which produce oocytes and hormones, and the reproductive tract, which consists of fallopian (or oviduct) tubes, uterus, and vagina, all of which are required for fertilization, pregnancy, and delivery of offspring. In addition to these primary sex organs, the hypothalamus and anterior pituitary play integral roles in maintaining homeostasis of the reproductive system by controlling the production of hormones. Any factor that interferes with the development and/or functions of these reproductive organs will have detrimental effects on the success of reproduction. The female reproductive system is susceptible to damages caused by various factors. These factors could be intrinsic, such as genetic disorders and/or physiological imbalances, or extrinsic, such as sexually transmitted diseases and/or man-made chemicals, or a combination of both. Exposure to harmful man-made or environmental chemicals has become a major concern in female reproductive health due to the fact that many chemicals have become an inseparable part of our daily life. Evaluating and understanding the potential toxic effects of a particular chemical on the female reproductive system is important for the development of not only preventive measurements but also treatment strategies if necessary. Various in vitro and in vivo approaches have been developed to determine the mechanism of action of a particular chemical. In vitro systems, such as cell, tissue, or organ culture, provide valuable insight into the direct effects of a chemical at the cellular level. The limitation of these in vitro systems is that often they are not able to address issues such as absorption, metabolism, distribution, and excretion of chemicals of interest. In vivo systems, such as dosing animals with chemicals, provide more realistic conditions to examine the systemic impact of chemicals; however, these systems are limited in their ability to identify the exact mechanisms of action. Furthermore, these genetically modified mice can be exposed to chemicals of interest to study changes in response to the exposure in comparison to normal wild-type animals. Understanding these genetic models is the first critical step to apply these models to toxicological research. These chemicals alter female reproduction either directly in their native form or indirectly via their metabolites. The native chemical or its metabolite, which structurally resembles an endogenous hormone, may bind to the corresponding hormone receptor and eventually antagonize or mimic the functions of endogenous hormones. Toxic effects via genomic pathways have been documented as a result of exposure to natural compounds. In mammals, development of the female reproductive system can be divided into two phases: hormoneindependent and hormone-dependent phases. The hormone-independent phase starts in embryonic life and extends to the prepubertal age. In mammals, fetal ovaries form as a result of the absence of the Y-linked testis-determining gene Sry (sexdetermining region of the Y chromosome) (Gubbay et al. Once the ovaries are formed, female germ cells or oogonia inside the ovaries progress through meiotic prophase, leptotene, zygotene, and pachytene stages, and are eventually arrested at the diplotene stage of the first meiotic division. At the same time, the somatic cells surrounding the oogonia give rise to granulosa cells and initiate the formation of primordial follicles. During this hormone-independent phase, sex steroids and hormones from the anterior pituitary are not required for the development of female sex organs. Instead, local growth factors and signaling molecules play major roles in molding the various parts of the female reproductive system. For example, when genistein is administered to pregnant mice between days 15 and 19 of pregnancy, this estrogenic compound accelerates the age of vaginal opening and causes irregular estrous cycles in the female offspring (Nikaido et al. The hormone-dependent phase of development of the female reproductive system starts at puberty when physiological, morphological, and behavioral changes occur. The ovaries start to release steroid hormones in response to gonadotropins from the anterior pituitary (Miller and Strauss, 1999). Production of sex steroids such as estrogens stimulates further differentiation of 474 Genetic Mouse Models for Female Reproductive Toxicology Studies the female reproductive tract, including vagina and uterus, and leads to the appearance of secondary sexual characteristics such as enlargement of the breasts in humans. In the ovary, the follicles undergo cyclical changes and differentiate from primordial follicles into primary, secondary, and antral follicles. Antral follicles are the only follicle type capable of releasing oocytes upon ovulation. They are also the major source of estrogens, which are required for inducing and maintaining menstrual/estrous cyclicity. As female animals age, the ovaries become devoid of follicles due to the processes of ovulation and follicular atresia. They can also target the ovaries and disrupt ovulation, increase rates of follicular atresia, and cause early onset of reproductive senescence. The phytoestrogen genistein is known to target reproductive organs including the uterus, causes spontaneous abortion prior to term, interferes with uterine development and function, alters folliculogenesis and steroidogenesis, and affects estrous cycles in rodents (Jefferson et al. Phthalates are chemicals used in the manufacturing of automotive parts, medical supplies, plastic wraps, and beverage containers. In rodents, phthalates have been shown to cause spontaneous abortions, birth defects, prolonged estrous cycle, suppressed or delayed ovulation, a reduction in the size of preovulatory follicles, and defects in folliculogenesis and steroidogenesis (Davis et al. In the ovaries, theca cells in follicles synthesize and secrete androgens, while granulosa cells convert these androgens to estrogens. Therefore, in the following section we focus specifically on transgenic models that affect steroidogenesis, hormone receptor functions, and hormone synthesis, and discuss potential applications of these genetic models to toxicological studies in the future. Genetic mouse models that develop phenotypes in the female reproductive system and potential chemicals of interest that could be tested in these models are listed in Table 1. Ovarian steroid hormones (progestins, androgens, and estrogens) are synthesized in a sequential manner by the theca and granulosa cells of follicles (Drummond et al. Several studies have used transgenic mouse models to examine the roles of genes in the steroidogenic pathway. Congenital adrenal hyperplasia has been also shown to have reproductive effects in female mice homozygous null for Cyp11b1 (Mullins et al. The female reproductive system is profoundly affected in null mice and displays endometrial hyperplasia and adenomyosis, presumably due to prolonged exposure to adrenal P4 (Mullins et al. Furthermore, female mice with the ovarian-specific deletion of Nr5a1 exhibited abnormal estrous cycles, were infertile, and displayed a blunted induction of E2 in response to gonadotropins (Pelusi et al. Female mice deficient for Nr5a1 model premature ovarian failure/primary ovarian insufficiency in humans (Harrison et al. Upon binding of ligands, nuclear receptors trigger the classical genomic pathway and work in concert with other transcription factors to regulate the expression of specific genes, thereby controlling the metabolism, development, and homeostasis of the cell. The actions of P4 are mediated via specific interactions with its corresponding receptors. Interestingly, E2 does not induce Pgr in the ovary, like it does in the uterus (Lydon et al. However, the inability to respond to P4 leaves these animals vulnerable to estrogenic insults. P4 also plays a role in the termination of sexual behavior during estrous cycle (Mani and Oyola, 2012). Mice become refractory to reproductive behavior upon further stimulation by the administration of P4 or E2 and P4, which is referred to as postestrous refractoriness (Mani and Oyola, 2012). Of particular importance to toxicological studies is that androgens can both promote and inhibit ovulatory function, which appears to be dose dependent (Walters, 2015).
Diseases
- Hyperglycinemia, isolated nonketotic type 2
- Apo A-I deficiency
- Bonnemann Meinecke syndrome
- Mediterranean fever[disambiguation needed]
- Adrenal incidentaloma
- TAU syndrome
- Tricho odonto onychodysplasia syndactyly dominant type
- Bone tumor (generic term)
- Acute myeloblastic leukemia type 4

Buy minipress line
Microtubules within growth cones are unstable and hiv infection graph 2.5bottles minipress overnight delivery, therefore, constantly experience periods of catastrophic disassembly and rescue. As this occurs, the microtubules grow and retreat from the actin-rich peripheral regions of the growth cone. Tau is thought to be an important factor in neurite development since inhibition of tau synthesis arrests neurite formation in cultured neurons (Caceres et al. The mechanism of neurite growth and guidance is based on the ability of the actin-rich filopodia and lamellipodia to recognize and react to extrinsic signals. Although guidance of growth cones is dependent upon actin dynamics, neurite extension can occur in a microtubule-dependent fashion in the absence of actin polymerization. When a microtubule grows into a filopodia that has been activated by an extrinsic signal, the microtubule is captured and stabilized. Organelles are then transported along the newly stabilized microtubule and a new section of neurite is established. This process allows vectoral growth of the neurites in the direction of, or away from, certain extracellular signals (Gordon-Weeks, 1993). Extension of lamellipodia and filopodia in growth cones occurs due to polymerization at the plus ends of actin filaments, which are oriented towards the membrane, and can push out on the membrane. Therefore, the rate of extension relies on relative rates of actin polymerization versus retrograde flow (Gomez and Letourneau, 2014). Guidance cues signal through pathways that regulate actin-binding proteins in specific locations within growth cones, and therefore determine sites and rates of polymerization/depolymerization. Cofilin activity is primarily regulated by phosphorylation, being in the active form when not phosphorylated (Bernstein and Bamburg, 2010). Intermediate filaments are not present in growth cones, but are essential for neurite development and circumferential axonal growth. Vimentin, a-internexin, and nestin are expressed during early stages of development and their disappearance coincides with the appearance of neurofilament subunits (Shea and Nixon, 1988). Inhibition of vimentin synthesis blocks neurite outgrowth in cultured cells, implying that intermediate filaments play a crucial, early role in neurite formation (Shea et al. Dendritic spines are supported by F-actin, and actin dynamics are essential for experience-dependent alterations to spines and synapses (Spence and Soderling, 2015). Many of the same actin regulators essential for growth cone motility are also involved in activity-dependent actin remodeling in spines. For example, Arp2/3 and formins are essential for spine maintenance and morphological alterations, profilin recruitment to spines is activity-dependent, and changes in cofilin activity are required for spine plasticity (Spence and Soderling, 2015). Because of the major regulatory role that actin plays at synapses, it is no surprise that actin dynamics are essential for learning and memory (Lamprecht, 2014). Following several learning paradigms, it was found that disruption of actin dynamics by cytochalasin D or latrunculin A inhibits memory formation in rats. For example, cofilin phosphorylation has been shown to be increased after training on several learning tasks, and knock-down of cofilin results in impaired memory (Rust et al. Freeze-fracture, freeze-etch, and quickfreeze preparations for electron microscopy reveal thin, filamentous attachments between the different cytoskeletal proteins (Hirokawa, 1982; Hirokawa et al. This suggests that the interconnections are dynamic and that the filamentous components of the cytoskeleton may communicate through conformational rearrangement, a contention further supported by the observation that under certain conditions, disruption of microfilaments or microtubules reversibly reorganizes the intermediate filament network (Bershadsky and Vasiliev, 1988; Tint et al. The interplay between microtubule and actin dynamics is especially important for neurite growth and stability, as well as neuronal migration. In growth cones, actin and microtubule assembly/disassembly and localization involve important interactions (Coles and Bradke, 2015). Interactions between microtubules and actin are regulated by several proteins that can bind to both microtubules and actin (Coles and Bradke, 2015). The signal pathways that lead from extracellular signals to actin reorganization have been intensively studied. The preceding overview of the cytoskeleton underscores the complexity of these organelles and their interactions; this complexity gives rise to a correspondingly diverse spectrum of toxicant effects. Toxin interactions with the major cytoskeletal proteins, associated proteins, or regulatory proteins may cause impairment of function and cell death. Moreover, differences in cytoskeletal function between developing and mature neurons contribute to the differential sensitivity to toxicant effects in the fetal and adult nervous systems. The dynamic nature of cytoskeletal structures and their sensitivity to alterations in the cellular microenvironment make them highly vulnerable to toxins, often with adverse effects on cellular function. Moreover, because of the structural and functional linkages between cytoskeletal elements, attack upon one component may result in secondary changes to other components. It is of interest to note that much of the work describing the normal dynamics and function of the cytoskeleton has been accomplished using neurotoxins as tools; many unwanted side effects of therapeutic agents and toxicities of environmental chemicals are mediated via similar mechanisms critically involving the cytoskeleton. This can be accomplished by disrupting the microtubules of the mitotic spindle apparatus. The mechanisms of microtubule disruption vary from direct polymer stabilization to inhibition of polymerization through alterations in microtubule regulation (Negi et al. Unfortunately, antineoplastic compounds react with all microtubules, including those in neurons, and produce characteristic axonopathies (Gupta and Abou-Donia, 1994). Axonopathies usually develop following relatively high doses, whereas mitotic spindle formation is inhibited at much lower doses, which makes these compounds useful in cancer treatment. A wide range of natural products have been, and are being, studied for their anticancer activities. Not all of these products are being used in human trials so it is not possible to assess whether they will cause neuropathies or axonopathies Cytoskeletal Elements in Neurotoxicity 107 in patients, but the relationships among products being studied and those currently in use indicates that the possibility of concomitant neurotoxicity should be expected. These products will be discussed in this section, along with those that are known to cause neuropathies. Vinblastine and vincristine, antineoplastic vinca alkaloids, were the first natural products used for chemotherapy. These compounds, and their analogs, bind free tubulin and prevent tubulin aggregation and polymer stabilization (Kingston, 2009). In treated cells free tubulin becomes bound in nontubular aggregates, the concentration of tubulin is decreased, and microtubule polymerization is inhibited. At low concentrations vinblastine stabilizes polymerized microtubules in vitro and inhibits dynamic instability (Toso et al. Combretastatin, a related inhibitor of tubulin polymerization from the root of Combretum caffrum, is more cytotoxic than the vinca alkaloids (Kingston, 2009). The combretastatin compounds, and their analogues, are being developed for clinical trials. Dolastatin, isolated from the sea hare Dolabella auricularia (see discussion under Biological Agents), affects both microtubules and microfilaments. This compound, and its analogs, bind tubulin and inhibit polymerization (Kingston, 2009). In clinical trials 40% of patients develop some level of peripheral neuropathy, which limits the usefulness of this compound for chemotherapy. Nocodazole and oncodazole, chemotherapeutic benzimidazole derivatives, are similar in structure to the vinca alkaloids but bind with high specificity to the plus ends of microtubules. Nocodazole enters cells, binds to unpolymerized tubulin, and blocks microtubule assembly (Baas and Black, 1990; Baas et al. This binding results in a net loss of microtubules from the axon or dendrite due to continued disassembly of microtubules without concomitant assembly. Nocodazole reduced formation of dendrites in primary cultures of hippocampal neurons (Witte et al. In the same study low doses of taxol induced formation of extra axons in neurons with sprouting neurites. Taxol (Paclitaxel), an antineoplastic compound isolated from the Pacific yew, binds polymerized microtubules, stabilizes their structure, and causes the slow accumulation of microtubule bundles in the cytoplasm. There is an overall decrease in the rate of microtubule polymerization caused by a decrease in the concentration of free tubulin. Unlike the vinca alkaloids that sequester free tubulin, the decreased concentration of tubulin occurs because of a decrease in the rate of depolymerization, which would normally add new dimers to the pool of free tubulin. Taxol appears to have no effect on the rate of fast axonal transport, but an inhibition of retrograde axonal transport has been reported (Nennesmo and Reinholt, 1988). Since the motors for anterograde and retrograde fast transport both move along microtubules, it is interesting to speculate that taxol may interfere with dynein binding or function. Taxol also causes an almost complete inhibition in slow anterograde tubulin transport (Komiya and Tashiro, 1988). This is not solely due to the decrease in free tubulin since the movement of actin and other proteins in slow component b is also blocked.
Order 1 mg minipress amex
Future work could also examine ovarian metabolism of chemicals using this system xl3 accion antiviral order minipress 2.5mg fast delivery, separating activity of the ovary from other tissues, especially the liver. Most culture methods for intact follicles were designed to ensure that follicles kept their spherical structure with an intact basal membrane throughout the culture. Therefore, follicles needed to be supported by an artificial extracellular matrix or prevented from attaching to the culture vessel by different means. These cultures are successful in terms of steroid production and oocyte maturation; however, they only sustain follicle growth from the antral stage, and microscopic observation of follicle growth and development during culture is difficult and hard to quantify (Demeestere et al. In the culture method that allows follicles to attach to the bottom of the culture vessel, follicles remodel to open 3D structures, while keeping their cell proliferation and differentiation pattern close to the in vivo ovarian physiology (Cortvrindt and Smitz, 1998). Conversely, follicles growth in a 3D culture system have been shown to more closely resemble the in vivo environment by also providing optimal physical parameters such as reproducing the rigidity of the ovarian stroma (Shea et al. Steroid hormone (androstenedione, testosterone, E2, and P4) and growth factor (inhibins, activin) concentrations can be measured in the spent medium. The developmental competence of the oocytes was proven by in vitro fertilization and embryo transfer giving rise to live young (Liu et al. Day 12: Preovulatory follicle: large antral follicle with clearly visible antral-like cavity (A). Day 13: Upon an ovulatory stimulus, the central oocyte cumulus complex is free floating in the culture droplet. Adaptation of the culture method to an oil-free environment gave it the potential to be used as an in vitro bioassay to assess the impact of a variety of compounds on the ovarian function in vitro (Cortvrindt and Smitz, 2002) and be part of an in vitro battery to replace, reduce, or refine the in vivo fertility study for reproductive toxicity assessment of chemicals (Bremer et al. For this purpose, the follicle culture method was highly standardized and proven to be reproducible (Table 2). Relevant morphological and biochemical evaluation parameters for folliculogenesis, oogenesis, and steroidogenesis allow pinpointing the influence of compounds on the ovarian function in vitro. The integration of these different outcome parameters provides information on the well-being of female reproductive health. If a compound has the ability to affect ovarian function, it will be recognized by deviation of the normal values of the selected end point parameters: interruption of folliculogenesis is an indication of menstrual cycle disturbances; alteration of the steroid output points to cycle disturbances, ovulation problems, alterations of the uterine receptivity; and aberrant oocyte development points to impairment of conception or the 526 In Vitro Ovarian Model Systems risk of early abortion (aneuploidies). In addition, detailed analysis of the steroid output at different days gives information on the enzymatic activity of all the enzymes involved in the steroid pathway and can identify endocrine disruptors. Various matrixes have been characterized but alginate-based matrices seem to be the most common for isolated follicles thus far. Various studies have described a fibrin alginate that more closely resembles the follicular microenvironment in terms of topology and mechanical stimulation and that, when used with rodent follicles, shows great promise in high-throughput toxicity screening (Zhou et al. One key feature of this system is the ability to culture follicles starting from the primordial stage. Primordial follicles are isolated from the ovaries of various species and encapsulated in high hydrogel concentrations. By using high concentrations, this environment mimics the dense cortex characteristics of the in vivo ovarian environment where primordial follicles reside (Shea et al. Specifically, primordial follicles are encapsulated by pipetting primordial follicle-containing 0. Several studies applying test compounds belonging to different chemical classes have been performed to prove the usefulness of the bioassay in the domain of reproductive toxicology. Furthermore, the coupling of this bioassay with novel techniques such as adenoviral gene expression systems will further enhance the contribution of this in vitro system to mechanistic toxicology. Some benzodiazepines like diazepam bind to both types of receptors; others, on the contrary, exert their influence in a selective way. It is well known that anticancer agents have a detrimental impact on ovarian function. Follicles were exposed throughout the culture to flumazenil, diazepam, or chlorodiazepam at the respective concentrations. Flumazenil had no impact on follicle development or survival, whereas chlorodiazepam and diazepam concentration-dependently reduced follicle survival and development. Flumazenil had no impact on oocyte maturation rate, whereas diazepam and chlorodiazepam significantly reduced oocyte maturation capacity. Dexamethasone dramatically disturbed E2 and testosterone secretion patterns at all doses tested. Secretion of E2 was decreased by more than 65% on day 8 and 12 of culture, as compared to controls. On day 12, testosterone levels were up to 85% lower in all treatment groups than in the control. The decreased E2, testosterone, and P4 levels confirm the direct effect of dexamethasone induced by downregulation of steroidogenic acute regulatory protein expression, as previously shown in rats (Huang and Shirley Li, 2001). Although it is generally assumed that antibiotics are quite harmless to eukaryotic cells, at present, only a few studies are available that analyze the potential reproductive toxicological effects. Penicillin is a b-lactam antibiotic that acts on Gram-positive bacteria by inhibiting the formation of the bacterial cell wall and thereby preventing replication. Even at the highest concentration tested, no effect was detected at the level of follicle survival or differentiation; also oocyte development and maturation were completely normal at all tested doses. Streptomycin and gentamicin both belong to the class of aminoglycoside antibiotics. Streptomycin acts through inhibition of protein synthesis at the level of the prokaryotic ribosome (Luzzatto et al. This exposure did not affect follicle survival, but did impair follicle differentiation. These results demonstrated that long-term exposure to penicillin poses no problems to oogenesis while the aminoglycoside antibiotics streptomycin and gentamicin affect oocyte quality, indicating that caution should be taken in using these antibiotics in cell culture media for assisted reproductive technology. Follicle survival was not affected by the compound, but follicle differentiation (antrum cavity formation) was significantly accelerated in a dose-dependent manner. The soy isoflavone genistein is a tyrosine kinase inhibitor with estrogenic activity affecting ovarian function (Jefferson et al. It is the most toxic member of a class of chemicals called dioxins, which persist as contaminants in the human food chain, fresh water sources, and as aerosols due to industrial waste (Centers for Disease Control and Prevention, 2005). It is well documented that the AhR mediates some of the toxic effects of endocrine-disrupting chemicals and that global deletion of the AhR in female mice results in a reduced rate of growth of ovarian antral follicle (Hernandez-Ochoa et al.
Buy 2 mg minipress otc
Several federal agencies work cooperatively to identify and regulate population exposure to environmental chemicals antiviral definition purchase 2mg minipress mastercard, as well as drugs; note that drugs can also become effluents that enter the environment. The importance of the time dimension makes consideration of developmental toxicology in risk assessment a daunting undertaking. The period for disruption of normal human development begins at conception and extends through puberty and into early adulthood until all developmental processes cease. The protracted timeline is punctuated with multiple time windows of particular sensitivity for many different organs and biological systems. Consequently, there exist countless developmental toxicology mechanisms, many barely known, and few well understood. What we can guess is that a finite number of genes and biological pathways are involved in the expression of a particular developmental toxicity. With the advent of inexpensive high-throughput assays, together with sequencing data, computational approaches offer a means to begin to unravel the complex high-dimensional molecular processes of developmental toxicity pathways. As toxicity expression pathways are elucidated, the potential exists to refine assays and develop new ones that, in turn, can be expected to yield larger and more diverse predictive model training data sets, for both currently known developmental toxicology endpoints and those to be discovered. Thus, while a specific toxicity mode of action may manifest differently across development, if an agent is active in that mode, at least we are aware of a need to look at a particular development stage. Developmental disturbances are complicated by differing exposure and metabolism profiles for the fetus and mother. In most cases, in utero and postnatal exposures throughout development and Absorption, Distribution, Metabolism, and Excretion properties for chemicals can only be crudely estimated (Ladics et al. Thus, animal testing is and will be continuously a de facto approach for developmental toxicity, an approach presumed cautionary has long been used. Specifically, human risks are extrapolated from the animal in vivo experiments as dictated by law, with limits to human exposure historically set by applying an additional cross-species safety factor to the no-observed-effects-level (Olson et al. The appropriateness of rules and reliance on the data from such an old paradigm is perpetually in a debate (Knight, 2007; Bailey et al. Testing toxicity in animal models is a protracted and expensive process consuming many animals. Most unfortunately, in vivo test results from animal models are not necessarily reproducible and even relevant to humans due to the difficulty of controlling experimental variability, which add the challenge of correlating its findings with human toxicity (Rhomberg, 2010). These concerns in developmental toxicity testing are not new, and the methodologies prescribed by law are timeworn. Even though in place since the 1970s, the types and amounts of data relevant for developmental toxicology are broadly inadequate, and what data there are cover but a small portion of the chemical universe. Thus, computational toxicology approaches such as read-across may have a role in this field, a topic that is extensively discussed in the following sections. The previous version was also quite prescient in laying out a framework for the future; as that framework has proven valid, we include the last addition in its entirety by reference (Singh et al. Together, the previous issue and this one delve into key elements of the ongoing paradigm shift in computational toxicology. Specifically, in this article, we focus our discussion of computational toxicology as a part of a paradigm shift in science. While the previous edition was written through the lens of a biologist, we write here through the lens of a computational toxicologist. The views between the last edition and this one are highly complementary, given that computational toxicology is inherently multidisciplinary. We have already declared that the computational toxicology comprises the building of models of many different types. Much of the article will be devoted to model building, a topic not covered in the previous edition. Mathematics, bioinformatics, statistics, machine learning, and computational sciences use different terminologies. The article is divided into four broad areas: (1) computational toxicology resources, (2) computational toxicology projects, (3) computational toxicology modeling, and (4) future directions and perspective. We conjecture that integrative modeling approaches, which can be addressed in many ways, will emerge to prominence across computational toxicology. An integrative approach using, for example, biological pathways to inform feature selection for models and biomarkers will yield both greater accuracy and a better understanding of modes of action. In the second section, we summarized some large projects, most from international collaborative efforts, that contribute to the significant advancement as we have seen nowadays in computational toxicology. Along this line, ontology (and controlled vocabularies) to support computational toxicology will be discussed with some examples. The remainder of the article is devoted to modeling, starting with describing the mathematical logic in building valid models, then the enormous flexibility in methods available, model validation, and the increasingly important topic of model reliability. The last part of the modeling discusses text mining by an approach not included in the prior edition, generative models that require no data labeling; these models perform both dimension reduction and data labeling according to latent variables. Latent modeling approaches allow documents in a large unstructured corpus to be rapidly structured and rank ordered in agreement with the probability of relatedness to a query. They are also effective means for obtaining the equivalent of clustering analysis, allowing any type of data to be integrated. We conclude, as Future Directions and Perspective, with a summary of how computational science will be the indispensable cornerstone in future advances in biological, medical, and toxicological sciences. Opportunities for replacing most expensive, slow, and nonpredictive animal and in vivo testing with predictive modeling are ubiquitous. Criticism has been mounting in high impact journals that much if not most research findings are irreproducible, and that a fundamental tenet of science, reproducibility, has been forsaken in the name of competition for novel positive outcomes. Risks of false positive and false discoveries are inherently high in biology due to uncertainty of causal variables. The future demands that both methods and model validity and reliability have to be carefully vetted. Increasingly, large collaborative teams are forming to carry out an immense work required to define the best approach in a complex from data collection, to interpretation to final model. Finally, as any computational model or predictor can, by definition, be no better than the data it is based on, experimental protocols and models need to move toward symbiotic codependency. Examples are disease type, toxicity type, a chemical structure-dependent molecular mechanism, or some combination of them (Ding et al. The data could range from being unstructured to being structured and possibly highly structured in an ontology framework. Beyond the value of aggregated high-quality data per se are the abilities to build models and make inferences. Better still, data points will be carefully selected to span the n-dimensional space in which models will be developed and validated. Additionally, an ensemble of models could be built to form a single integrative predictor. Ensembles offer the potential of combining many weak signals into a stronger one, enhancing both reliability and validity. Of course, there are many algorithms, both supervised and unsupervised, that can be employed (Maggioli et al. With the advent of inexpensive high-throughput sequences data, integrative modeling approaches combining old data with new information are anticipated to become more prevalent. Endocrine disruptor compounds are mostly the environmental chemicals that have the ability to disrupt the endocrine systems of animals and humans. Bioactivity is exerted through transcription factors formed with nuclear receptor superfamily dimers (that are highly conserved across vertebrates). Endocrine disruptors produce a harmful effect on male and female reproductive system as well as the development of the fetus. Modern human environments and life forms in environmental ecosystems are exposed to tens of thousands of chemicals for which little to no safety testing data exists. A significant number of the environmental chemicals are persistent in nature, and thus bioaccumulation is seen up the food chain (Kraus et al. Only recently (Tollefson, 2016) has legislation been passed that empowers regulators to require safety testing of tens of thousands of environmental chemicals. With some 85,000 chemicals, of which only about 2% have been reviewed for safety, computational toxicology combined with high-throughput assays will be essential (Tollefson, 2016). So, here again, the new paradigm using a combination of modern sequencing and high-throughput assays to detect activity in toxicity related pathways appears to be the viable way forward.
Generic 2.5bottles minipress
Fumonisins are present at low levels in maize throughout the world antiviral for cold order 1mg minipress with mastercard, but high levels of the toxin may occur, depending on the environmental conditions and genetics of the host plant. Strategies for reducing the risk of fumonisin contamination in maize supplied to the market include improved crop management practices, improved breeding strategies, as well as transgenic approaches for increased ear-mold resistance (Betz et al. Oxidative damage and fumonisin B1-induced toxicity in primary rat hepatocytes and rat liver in vivo. Global birth prevalence of spina bifida by folic acid fortification status: A systematic review and meta-analysis. Expression patterns of folate binding proteins one and two in the developing mouse embryo. Temporal expression of fumonisin B(1)-induced tumor necrosis factor-alpha and interferon gamma in mice. The kidney in vitamin B12 and folate homeostasis: Characterization of receptors for tubular uptake of vitamins and carrier proteins. Megalin is essential for renal proximal tubule reabsorption and accumulation of transcobalamin-B(12). Neural tube defects in Australia: Trends in encephaloceles and other neural tube defects before and after promotion of folic acid supplementation and voluntary food fortification. Sphingosine 1-phosphate receptors in health and disease: Mechanistic insights from gene deletion studies and reverse pharmacology. Concentrations of blood folate in Brazilian studies prior to and after fortification of wheat and cornmeal (maize flour) with folic acid: A review. Effect of micronutrient fortification on nutritional and other properties of nixtamal tortillas. Gangliosides of anencephalic and fetal braindimmunostaining on thin-layer chromatograms. Neural tube defects: A review of human and animal studies on the etiology of neural tube defects. Amelioration of intracellular stress and reduction of neural tube defects in embryos of diabetic mice by phytochemical quercetin. Oxidant regulation of gene expression and neural tube development: Insights gained from diabetic pregnancy on molecular causes of neural tube defects. The Costa Rican experience: Reduction of neural tube defects following food fortification programs. Functional variant in methionine synthase reductase intron-1 is associated with pleiotropic congenital malformations. Fumonisin B1, a mycotoxin contaminant of cereal grains, and inducer of apoptosis via the tumour necrosis factor pathway and caspase activation. Fumonisin, Folate and other Methyl Donors and Neural Tube Defects 195 Cifuentes, G. Perfil epidemiologico de law anomalias del tubo neural en Guatemala, durante el ano 2000. Graduation Thesis of the School of Medicine of Universidad San Carlos de Guatemala. Whole rat embryos require methionine for neural tube closure when cultured on cow serum. Wheat flour fortification with folic acid: Changes in neural tube defects rates in Chile. Chemokines, sphingosine-1-phosphate, and cell migration in secondary lymphoid organs. Prevention of the first occurrence of neural-tube defects by periconceptional vitamin supplementation. Antibodies to folate receptors impair embryogenesis and fetal development in the rat. Effect of nixtamalization (Alkaline cooking) on fumonisin-contaminated corn for production of masa and tortillas. The Nrf2 activator vinylsulfone reduces high glucose-induced neural tube defects by suppressing cellular stress and apoptosis. Fumonisin B1 affects viability and alters nitric oxide production of a murine macrophage cell line. Prospects for reducing fumonisin contamination of maize through genetic modification. Methionine but not folinic acid or vitamin B-12 alters the frequency of neural tube defects in axd mutant mice. Epigenetic alterations in folate transport genes in placental tissue from fetuses with neural tube defects and in leukocytes from subjects with hyperhomocysteinemia. Perturbations in choline metabolism cause neural tube defects in mouse embryos in vitro. Assessment of the embryotoxic potential of the total hydrolysis product of fumonisin B1 using cultured organogenesisstaged rat embryos. Unbiased quantitative proteomics of lipid rafts reveals high specificity for signaling factors. Elevated nuclear sphingoid base-1-phosphates and decreased histone deacetylase activity after fumonisin B1 treatment in mouse embryonic fibroblasts. The cancer-promoting potential of fumonisin B1 in rat liver using diethylnitrosamine as a cancer initiator. Fumonisin-induced hepatocarcinogenesis: Mechanisms related to cancer initiation and promotion. Maternal fumonisin exposure and risk for neural tube defects: Mechanisms in an in vivo mouse model. Embryonic development in the reduced folate carrier knockout mouse is modulated by maternal folate supplementation. The characteristics of the membrane transport of amethopterin and the naturally occurring folates. The mycotoxin fumonisin B1 transiently activates nuclear factor-kappaB, tumor necrosis factor alpha and caspase 3 via protein kinase C alpha-dependent pathway in porcine renal epithelial cells. Modelling fortification of corn masa flour with folic acid and the potential impact on Mexican-American women with lower acculturation. Oxidative stress is implicated in arsenic-induced neural tube defects in chick embryos. Antisense modulation of the coding or regulatory sequence of the folate receptor (folate binding protein-1) in mouse embryos leads to neural tube defects. Rat kidney pathology induced by chronic exposure to fumonisin B1 includes rare variants of renal tubule tumor. Fumonisin toxicosis in swine: An overview of porcine pulmonary edema and current perspectives. Signaling through sphingolipid microdomains of the plasma membrane: the concept of signaling platform. Interaction of 5-methyltetrahydrofolate and tetrahydrobiopterin on endothelial function. Microdomain-dependent regulation of Lck and Fyn protein-tyrosine kinases in T lymphocyte plasma membranes. Characterization of the human and mouse sphingosine 1-phosphate receptor, S1P5 (Edg-8): Structure-activity relationship of sphingosine1-phosphate receptors. Transfer of folic acid inside the first-trimester gestational sac and the effect of maternal smoking.
