

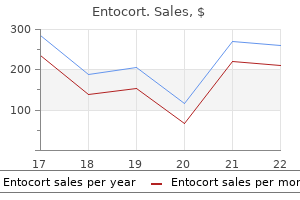
Purchase entocort 100mcg on-line
The neural correlates of emotion-based cognitive control in adults with early childhood behavioral inhibition allergy forecast vero beach fl cheap entocort online american express. Differential effects of social and nonsocial reward on response inhibition in children and adolescents. Neurophysiological correlates of executive function: a comparison of EuropeanCanadian and Chinese-Canadian 5-year-old children. Temperamental exuberance and executive function predict propensity for risk-taking in childhood. Early behavioral inhibition and increased error monitoring predict later social phobia symptoms in childhood. Cognitive conflict links behavioral inhibition and social problem solving during social exclusion in childhood. Neural activation underlying cognitive control in the context of neutral and affectively charged pictures in children. Incentive motivation, cognitive control, and the adolescent brain: is it time for a paradigm shift Adolescent Brain Development: Implications for the Juvenile Criminal Justice System. Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Event-related functional magnetic resonance imaging of reward-related brain circuitry in children and adolescents. A history of childhood behavioral inhibition and enhanced response monitoring in adolescence are linked to clinical anxiety. The neural correlates of cognitive control and the development of social behavior Chapter 24 537 Meyer, A. Annual Research Review: On the relations among self-regulation, self-control, executive functioning, effortful control, cognitive control, impulsivity, risk-taking, and inhibition for developmental psychopathology. Development of affective decision-making for self and other: evidence for the integration of first- and third-person perspectives. Behavioral approach-inhibition in toddlers: prediction from infancy, positive and negative affective components, and relations with behavior problems. Progressive increase of frontostriatal brain activation from childhood to adulthood during event-related tasks of cognitive control. Linear age-correlated functional development of right inferior fronto-striato-cerebellar networks during response inhibition and anterior cingulate during error-related processes. The development of executive functioning and theory of mind: a comparison of Chinese and U. Event-related potentials to auditory stimuli following transient shifts of spatial attention in a Go/Nogo task. The expected value of control: an integrative theory of anterior cingulate cortex function. Effortful control, executive attention, and emotional regulation in 7e10-year-old children. Anterior cingulate cortex responds differentially to expectancy violation and social rejection. Neurophysiological mechanisms of emotion regulation for subtypes of externalizing children. Performance monitoring in children and adolescents: a review of developmental changes in the error-related negativity and brain maturation. Error-related brain activity in young children: associations with parental anxiety and child temperamental negative emotionality. Behavioral inhibition and anxiety: the moderating roles of inhibitory control and attention shifting. Chapter 25 Executive function: development, individual differences and clinical insights Hughes Claire1, 2 1 Newnham College, Cambridge University, Cambridge, United Kingdom; 2Centre for Family Research, Cambridge University, Cambridge, United Kingdom Chapter outline 25. Normative developmental trajectories for executive function from infancy to adolescence 540 25. From biological to environmental predictors of individual differences in executive function 545 25. Early executive function predicts academic, sociocognitive and social success at school 548 25. As Tau and Peterson (2010) put it, documenting normative pathways of brain development "provides the Archimedean point from which to interpret and understand the aberrant pathways of brain development that produce disease," while studying atypical groups "informs our knowledge of normal brain development by throwing into relief the developmental pathways that are most sensitive to perturbation. To address this gap, the first section of this chapter provides a synopsis of research findings on developmental trajectories from infancy to adolescence, in both typical and atypical populations. However, most studies in this field have been cross-sectional in design and have not controlled for peripheral task demands. These notes of caution need to be borne in mind when considering the summary review of findings from infants to adolescents given in the following. Early studies indicated that while eight-month-olds and older babies typically search correctly at location B, 5-month olds persist in searching for the object at location A. Although Piaget interpreted this "A-not B" error as a failure to recognize that objects have an independent existence in the world. Thus, young babies who make the A-not B error in their reaching responses may know that the object has been moved but fail to inhibit their previously successful (and therefore prepotent) reach to A. Other tasks that also demonstrate early executive skills in infancy include a detour-reaching task, in which infants are invited to retrieve an object that is visible behind a Perspex screen; success on this task depends on making a "detour reach" around the side of the screen (Diamond et al. In short, a rudimentary ability to inhibit prepotent responses is clearly evident by 7e12 months (Diamond, 2002). Together, these two studies indicate that executive attention (indexed by anticipatory looking) is also evident very early in life. Another reason for a research focus on preschoolers is that a wide variety of age-appropriate tasks have been developed in response to the previous dearth of tasks suitable for young children. Although it is difficult to summarize briefly, three points deserve particular mention. Specifically, investigations that straddle early infancy to toddlerhood (Cuevas and Bell, 2014; Devine et al. Latent variable analyses have also been used to conduct person-centered analyses. Specifically, as noted by Asbury and Plomin (2013), genetic influences are typically diffuse rather than specific and underpin stability rather than change over time. Specifically, while (as noted earlier) the preschool years are characterized by dramatic improvements in inhibitory control, evidence from young school-aged children and preadolescents highlight, respectively, improvements in cognitive flexibility and in working memory/planning ability. Luciana (2003) also showed that clear improvements on the Tower of London planning task or on a self-ordered search test of working memory were often not evident before the age of 11 or 12 years. For example, several studies report improvements in mental flexibility around age 8 years. At odds with this general pattern, however, are findings from another study that suggest a long developmental progression for cognitive flexibility, with 13-year-olds still not at adult levels (Davidson et al. According to this theory, the brain and mind work effortlessly (or under difficult conditions) at a gross level, but require effort (or more optimal conditions), to work in a more selective manner. Thus, it is easier to inhibit a dominant response all the time than only some of the time. As a result, even older children are likely to show frequent errors on task-switching paradigms, such as that used in Davidson et al. For example, adults slowed down on difficult trials to preserve accuracy, but children (and especially young children) were impulsive and so made errors on difficult trials. Here, one task battery that deserves mention is the behavioral assessment of dysexecutive syndrome (Roy et al. In a cross-cultural study to adopt this approach with a sample of 1427 participants (children aged 9e16 and their parents), Ellefson et al. Similar conclusions emerge from a community-based study of 10- to 12-year-olds in which self-reported impulsivity was inversely associated with both working memory and reversal learning (Romer et al. Arguing that sensation seeking is motivated by a desire to explore the environment in ambiguous risk contexts, these authors call for theoretical models that recognize adaptive roles of both cognition and experience during adolescence. In a metaanalysis of longitudinal studies involving children born very prematurely. The first of these was the "Windows" task in which, for each of 20 trials, children could win a treat (visible through a window in a box) by choosing a visibly empty box.
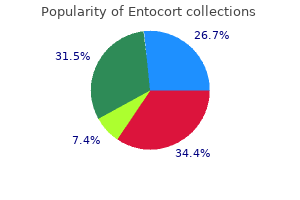
Order entocort online pills
Girls and women are better than boys and men in remembering the location of objects (Voyer et al allergy medicine heart patients 100 mcg entocort sale. Thus, it is not surprising that meta-analyses that combine across aspects of mathematics and ages disagree on the direction of the effect. Historically, the main sex difference was thought to be in problem-solving tasks, with boys outperforming girls, especially at older ages, but recent data show the sex difference to be small and to vary across nations. There is some indication that national differences are related gender equality across countries (Else-Quest et al. The sexes also differ in their variability in mathematical (and spatial) skills (Hedges and Nowell, 1995). Given the different patterns of sex differences in mathematics grades and standardized test scores, it is unclear the extent to which sex differences in mathematics may reflect other factors, such as anxiety. Indeed, mathematics anxiety has been shown to be greater for girls than boys and is a sex-specific predictor of performance (Devine et al. When there are differences, they are generally in the direction of girls and women having better skills than boys and men, although the size of the difference varies with age (Halpern, 2012). Language learning occurs earlier in girls than in boys, with boys catching up by age 6 years (Bornstein et al. In terms of specific verbal skills, females have a small to moderate advantage over males in several skills, most prominently in reading comprehension, verbal fluency, and phonological processing, but males have a small edge in analogies (Halpern, 2012). For instance, in the Programme for International Student Assessment, which includes data from 1. Boys write less well than girls, too, and this may be especially notable at the highest skill levels (Hedges and Nowell, 1995; Pargulski and Reynolds, 2017; Petersen, 2018; Wai et al. Girls and women also process verbal materials more rapidly than boys and men, including intelligence scale subtests, verbal fluency tests asking for many words to be generated quickly, letters of the alphabet, and digits (Camarata and Woodcock, 2006; Roivainen, 2011). But in several, girls and women are better than boys and men, with differences that are small to moderate in size: Compared with boys and men, girls and women show more accurate recall for learning facts or material that they read, more readily learn lists of words, have better recall for lists of common objects such as animals, food, furniture, and appliances, and have better recognition memory (Gur et al. The differences may be due in part to sex differences in strategy, with girls and women more likely to use clustering than boys and men. Sex differences in memory are especially consistent for episodic memory, that is, memory of specific events and episodes (Asperholm et al. For example, women are better than men at recognizing faces that they have seen before. Also, the types of events remembered seem to differ by sex, with women remembering more negative and less positive autobiographical experiences compared with men (Young et al. Many episodic memories are verbally based, and there is emerging evidence of heightened female superiority of this skill in cultures with high levels of female education and employment (Asperholm et al. Women also do better on episodic tasks, which require both verbal and visuospatial memory, but not when the tasks are clearly spatial, in which case, men do better (Herlitz et al. Importantly, these sex differences seem to be consistent across age (Herlitz and Rehnman, 2008). Moreover, research on working memory does not point to clear sex differences, perhaps because findings depend upon the stimuli to be remembered and task difficulty. In general, these tasks concern the ability to perceive details and shift attention quickly, often while using fine motor skills such as finger movements (Halpern, 2012). Boys and men typically identify as male, whereas girls and women typically identify as female (Blakemore et al. Sex differences in brain and behavioral development Chapter 27 591 As are most gendered phenomena, gender identity is multidimensional, with different aspects having different psychological antecedents, correlates, and consequences (Egan and Perry, 2001; Lippa, 2008), and there is recent work highlighting the value of considering degree of identification with both sexes (Martin et al. There are members of the population whose gender identity does not match their biological sex, including gender queer and nonconforming youth. There is considerable question and controversy over the nature and causes of transgender identity (see Korpaisarn and Safer, 2019; Polderman et al. For instance, gender dysphoria (distress) is difficult to determine, partly because it is highly dependent upon sampling and measurement. Moreover, rates of clinical referrals for gender dysphoria have historically been higher for males than females (Crissman et al. Most men and women have heterosexual orientation, with men being primarily attracted to women and women being primarily attracted to men (Blakemore et al. Not all individuals have exclusively heterosexual orientations or sexual experiences. There are sex differences in these rates, however, with women being more likely than men to consider themselves bisexual and to report changes in their sexual attraction and identities (Bailey et al. There is also evidence of sex differences in sexual arousal and acts regardless of sexual orientation. For instance, men show greater arousal to erotic images of their preferred sex. Moreover, men are more likely to engage in casual sex, masturbation, and pornography use (Petersen and Hyde, 2010). Given these sex differences in sexual behavior, some scholars have argued that sexual orientation has, to some degree, different determinants in women and men (Diamond, 2014). There are few sex differences in early milestones of reaching, sitting, crawling, and walking, but differences in motor skills begin to appear in the second year and continue through preschool (Koktejn et al. The earlier neurological development of s girls compared with boys results in early development of some skills, such as eyeehand coordination. Overall, girls develop sooner and are initially superior to boys with respect to fine motor skills, whereas boys tend to have greater strength and better gross motor skills (Flatters et al. There are, however, some exceptions to this, such as catching, in which there is evidence for a female advantage (Butterfield et al. Boys are also more physically active than girls in childhood and adolescence (Breslin et al. In adolescence, boys and girls continue to prefer and participate in different leisure activities. The main exception is effortful control, which shows a moderate advantage for girls. In addition, boys have higher levels of surgency than girls, perhaps reflecting their greater activity level. Research on "the big five" personality factors (neuroticism, extroversion, openness, agreeableness, and conscientiousness) shows that sex differences emerge in adolescence, leading to small, but relatively consistent, differences between adult men and women (De Bolle et al. It has been reported that women score higher than men in neuroticism (especially the anxiety and depression subscales), agreeableness, and extroversion (Costa et al. Cross-cultural similarities in the patterns of sex differences have also been found, but the differences may be largest in European and North American countries and smallest in African and Asian countries (Costa et al. There are also small sex differences favoring boys and men in global self-esteem, especially after childhood (Kling et al. Sex differences vary across aspects of self-esteem, with males having consistently higher appearance-related, personal self, self-satisfaction, and athletic selfesteem and females having higher ethical and behavioral conduct self-esteem (Gentile et al. Regarding emotional expression, there is some suggestion that women have more nonverbal emotion expression than men (Fischer and LaFrance, 2015) and that boys and men express more anger and less fear and sadness than girls and women; more convincing, however, is evidence that boys come to hide emotions such as sadness and fear purposefully, especially in the presence of peers (Chaplin, 2015; Kyratzis, 2001). Regarding perceiving emotion in others, girls and women are somewhat more sympathetic, empathic, and accurate at decoding emotions than are boys and men (ChristovMoore et al. Girls also show more interest in and have more nurturant interactions with babies than boys do, although the size of the difference varies across situations (summarized in Blakemore et al. Regarding risk taking, boys are more likely than girls to take risks and to be injured (Byrnes et al. Risk-taking behavior is relatively normative in adolescence (Steinberg, 2010), but the sex difference is generally seen in adulthood, as men report greater sensation seeking, disinhibition, boredom susceptibility, and adventure seeking than do women (Cross et al. Aggression is defined as behavior intended to harm others and includes physical, verbal, and indirect aggression. Although the social context affects aggression, and the size of the difference varies across age and culture, there are no instances in which females are more directly aggressive than males. Sometimes, physical aggression moves into the domain of seriously antisocial or criminal behavior. Although few people show the type of high levels of physical aggression that would be called violent or antisocial, or at the very extreme, commit murder, the majority of those who do are male, in childhood, adolescence, and adulthood (Archer, 2004; Dodge et al. Another form of aggression is called indirect, social, or relational aggression.
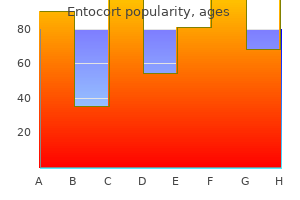
Cheap 200mcg entocort overnight delivery
Efferent and afferent connections of mouse sensory-motor cortex following cholinergic deafferentation at birth allergy testing auckland new zealand discount 100mcg entocort. Organization of cortical and thalamic input to pyramidal neurons in mouse motor cortex. Topographic precision in sensory and motor corticostriatal projections varies across cell type and cortical area. Frizzled3 is required for the development of multiple axon tracts in the mouse central nervous system. Transcription factor Lmo4 defines the shape of functional areas in developing cortices and regulates sensorimotor control. Cerebellar long-term depression: characterization, signal transduction, and functional roles. Projections of pyramidal tract cells to alpha-motoneurones innervating hind-limb muscles in the monkey. Comparative three-dimensional connectome map of motor cortical projections in the mouse brain. An anterograde tracer study of the developing corticospinal tract in the rat: three components. Distinct contributions of mesencephalic locomotor region nuclei to locomotor control in the freely behaving mouse. Corticospinal tract development and spinal cord innervation differ between cervical and lumbar targets. Integration of descending command systems for the generation of context-specific locomotor behaviors. Development of specificity in corticospinal connections by axon collaterals branching selectively into appropriate spinal targets. The primary motor cortex is involved in the control of a non-motor cognitive action. EphrinB3 is the midline barrier that prevents corticospinal tract axons from recrossing, allowing for unilateral motor control. Reevaluation of the primary motor cortex connections with the thalamus in primates. Two types of thalamocortical projections from the motor thalamic nuclei of the rat: a single neuron-tracing study using viral vectors. The development of motor control in the rhesus monkey: evidence concerning the role of corticomotoneuronal connections. Corticomotoneuronal synapses in the monkey: light microscopic localization upon motoneurons of intrinsic muscles of the hand. Satb2 regulates the differentiation of both callosal and subcerebral projection neurons in the developing cerebral cortex. The determination of projection neuron identity in the developing cerebral cortex. Cell biology in neuroscience: cellular and molecular mechanisms underlying axon formation, growth, and branching. Postnatal development of differential projections from the caudal and rostral motor cortex subregions. Generation of a whole-brain atlas for the cholinergic system and mesoscopic projectome analysis of basal forebrain cholinergic neurons. Areal distributions of cortical neurons projecting to different levels of the caudal brain stem and spinal cord in rats. Modulation of sustained electromyographic activity by single intracortical microstimuli: comparison of two forelimb motor cortical areas of the rat. Corticomotoneuronal connections in the rat: evidence from double-labeling of motoneurons and corticospinal axon arborizations. An experimental study of the cortico-spinal system in the monkey (Macaca mulatta). The spinal pathways and preterminal distribution of degenerating fibers following discrete lesions of the pre- and postcentral gyri and bulbar pyramid. Ryk-mediated Wnt repulsion regulates posterior-directed growth of corticospinal tract. Gene coregulation by Fezf2 selects neurotransmitter identity and connectivity of corticospinal neurons. Excitatory projection neuron subtypes control the distribution of local inhibitory interneurons in the cerebral cortex. Cerebral cortex assembly: generating and reprogramming projection neuron diversity. Tangential neuronal migration controls axon guidance: a role for neuregulin-1 in thalamocortical axon navigation. Robo1 and Robo2 cooperate to control the guidance of major axonal tracts in the mammalian forebrain. Corticospinal axons make direct synaptic connections with spinal motoneurons innervating forearm muscles early during postnatal development in the rat. Plastic changes in the vibrissa motor cortex in adult rats after output suppression in the homotopic cortex. Suppression of activity in the forelimb motor cortex temporarily enlarges forelimb representation in the homotopic cortex in adult rats. Long-range neuronal circuits underlying the interaction between sensory and motor cortex. Changes in the discharge patterns of cat motor cortex neurones during unexpected perturbations of on-going locomotion. Differential spinal projections from the forelimb areas of the rostral and caudal subregions of primary motor cortex in the cat. Harnessing activity-dependent plasticity to repair the damaged corticospinal tract in an animal model of cerebral palsy. The organization and collateralization of corticostriate neurones in the motor and sensory cortex of the rat brain. Tbr1 and Fezf2 regulate alternate corticofugal neuronal identities during neocortical development. Large-scale maintenance of dual projections by callosal and frontal cortical projection neurons in adult mice. Cortical reorganization of sensorimotor systems and the role of intracortical circuits after spinal cord injury. Mechanisms controlling the guidance of thalamocortical axons through the embryonic forebrain. Novel subtype-specific genes identify distinct subpopulations of callosal projection neurons. DeCoN: genome-wide analysis of in vivo transcriptional dynamics during pyramidal neuron fate selection in neocortex. Integration of posture and locomotion in acute decerebrate cats and in awake, freely moving cats. Cerebellar-induced locomotion: reticulospinal control of spinal rhythm generating mechanism in cats. Principles of diffusion tensor imaging and its applications to basic neuroscience research. The differential contributions of the parvocellular and the magnocellular subdivisions of the red nucleus to skilled reaching in the rat. Projection on the motor cortex of thalamic neurons with pallidal input in the monkey. Precentral cortical zones related to flexion and extension in two hindlimb movements in the monkey. Use-dependent alterations of movement representations in primary motor cortex of adult squirrel monkeys. A comparative neuroanatomical study of the red nucleus of the cat, macaque and human. Corticostriatal interactions during learning, memory processing, and decision making. Acetylcholine as a neuromodulator: cholinergic signaling shapes nervous system function and behavior. Intracellular potentials recorded from motoneurons following precentral gyrus stimulation in primate. The importance of combinatorial gene expression in early Mammalian thalamic patterning and thalamocortical axonal guidance.
Buy generic entocort pills
Neural systems for visual orienting and their relationships to spatial working memory allergy treatment ragweed 200mcg entocort otc. Object and spatial visual working memory activate separate neural systems in human cortex. Measures of frontal functioning and the emergence of inhibitory control processes at 10 months of age. Visuospatial dysfunction following unilateral brain damage: dissociations in hierarchical and hemispatial analysis. Do the right and left hemispheres attend to the same visuospatial information within a face in infancy Toward understanding commonalities in the development of object search, detour navigation, categorization, and speech perception. Behavioral and neural effects of chromatic isoluminance in the primate visual motion system. The influence of chromatic information on visual motion processing in the primate visual system. Infant motion: detection (M:D) ratios for chromatically defined and luminance-defined moving stimuli. A developmental study of covert orienting to peripheral visual cues covert visual orienting across the lifespan. Asymmetric development of dorsal and ventral attention networks in the human brain. Neural mechanisms involved in the processing of global and local aspects of hierarchically organized visual stimuli. Morphometric evaluation of the hydrocephalic brain: relationships with cognitive development. Spinal lesion level in spina bifida: a source of neural and cognitive heterogeneity. Attending to global versus local stimulus features modulates neural processing of low versus high spatial frequencies: an analysis with event-related brain potentials. Spatial frequency selection and integration of global and local information in visual processing: a selective review and tribute to Shlomo Bentin. A search for the origins of equivalence concepts through a microanalysis of block play. Developmental changes in cognitive and behavioural functioning of adolescents with fragile-X syndrome. The synchronization within and interaction between the default and dorsal attention networks in early infancy. Differential development of high-level visual cortex correlates with category-specific recognition memory. An evolving view of duplex vision: separate but interacting cortical pathways for perception and action. Development of large-scale functional networks from birth to adulthood: a guide to the neuroimaging literature. Aberrant parietal cortex developmental trajectories in girls with Turner syndrome and related visual-spatial cognitive development: a preliminary study. The fusiform face area subserves face perception, not generic within-category identification. Visual attention to global and local stimulus properties in 6-month-old infants: individual differences and event-related potentials. On the development of human face-processing abilities: evidence for hyperactivation of the extended face system in children. Hemispheric asymmetry in global/local processing: effects of stimulus position and spatial frequency. White matter microstructural abnormalities in children with spina bifida myelomeningocele and hydrocephalus: a diffusion tensor tractography study of the association pathways. Dissociation of object and spatial visual processing pathways in human extrastriate cortex. Distinguishing the functional roles of multiple regions in distributed neural systems for visual working memory. Visual-motor integration, visual perception and motor coordination in a population with Williams syndrome and in typically developing children. More is not always better: increased fractional anisotropy of superior longitudinal fasciculus associated with poor visuospatial abilities in Williams syndrome. Region-specific alterations in brain development in one- to threeyear-old boys with fragile X syndrome. Inhibition of return produced by covert shifts of visual-attention in 6-month-old infants. Understanding developmental changes in the stability and flexibility of spatial categories based on object relatedness. The functional development and integration of the dorsal and ventral visual pathways: a neurocomputational approach. Cortical networks for visual reaching: physiological and anatomical organization of frontal and parietal lobe arm regions. A comprehensive study of whole-brain functional connectivity in children and young adults. Progressive and regressive developmental changes in neural substrates for face processing: testing specific predictions of the Interactive Specialization account. Anomalous development of brain structure and function in spina bifida myelomeningocele. The fusiform face area: a module in human extrastriate cortex specialized for face perception. Human posterior parietal cortex maintains color, shape and motion in visual short-term memory. Effects of X-monosomy and X-linked imprinting on superior temporal gyrus morphology in Turner syndrome. Uniform connectedness and grouping in the perceptual organization of hierarchical patterns. Hemispheric differences are found in the identification, but not the detection, of low versus high spatial frequencies. Visual hemispheric asymmetries depend on which spatial-frequencies are task relevant. Two hierarchically organized neural systems for object information in human visual cortex. You can play 20 questions with nature and win: categorical versus coordinate spatial relations as a case study. Categorical versus coordinate spatial relations: computational analyses and computer simulations. Evidence for two types of spatial representations: hemispheric specialization for categorical and coordinate relations. The forest, the trees, and the leaves: differences of processing across development. Object individuation in 10-month-old infants: manipulating the amount of introduction. Functional neuroanatomy of visuospatial working memory in fragile X syndrome: relation to behavioral and molecular measures. Lateralization of categorical and coordinate spatial functions: a study of unilateral stroke patients. An early sex difference in the relation between mental rotation and object preference. Longitudinal development of human brain wiring continues from childhood into adulthood. Mental rotation performance in children with hydrocephalus both with and without spina bifida.

Purchase 200 mcg entocort overnight delivery
The neurons in most of the hippocampus also begin to connect early in development: synapses are present as early as 15 weeks gestational age allergy kingdom entocort 100 mcg online. Both the number and density of synapses increase rapidly after birth, and reach adult levels by approximately 6 postnatal months. Perhaps as a consequence, glucose utilization in the temporal cortex reaches adult levels at the same time. Thus, there are numerous indices of early maturity of major portions of the medial temporal components of the network. In contrast to early maturation of most of the hippocampus, development of the dentate gyrus of the hippocampus is protracted (Seress and Abraham, 2008). Indeed, neurogenesis in the dentate gyrus of the hippocampus continues throughout childhood and adulthood (Tanapat et al. It is not until 12 to 15 postnatal months that the morphology of the structure appears adultlike. Maximum density of synaptic connections in the dentate gyrus also is delayed, relative to that in the other regions of the hippocampus. In humans, synaptic density increases dramatically (to well above adult levels) beginning at 8 to 12 postnatal months and reaches its peak at 16e20 months. After a period of relative stability, excess synapses are pruned until adult levels are reached at about 4e5 years of age (Eckenhoff and Rakic, 1991). Although the functional significance of later development of the dentate gyrus is not clear, there is reason to speculate that it impacts behavior. As already noted, upon experience of an event, information from distributed regions of cortex converges on the entorhinal cortex. From there, it travels to the hippocampus in one of two ways: via a "long route" or a "short route. This implies that maturation of the dentate gyrus of the hippocampus may be a rate limiting variable in the development of episodic memory early in life. Finally, hippocampal volume continues to increase gradually throughout childhood and into adolescence. Myelination in the hippocampal region continues throughout adolescence (Arnold and Trojanowski, 1996; Benes et al. The association areas also undergo a protracted course of development (Bachevalier, 2014). For example, it is not until the seventh prenatal month that all six cortical layers are apparent. The density of synapses in prefrontal cortex increases dramatically at eight postnatal months and peaks between 15 and 24 months. Pruning to adult levels does not begin until late childhood; adult levels are not reached until late adolescence or early adulthood (Huttenlocher, 1979; Huttenlocher and Dabholkar, 1997). In the years between, in some cortical layers there are changes in the size of cells and the lengths and branching of dendrites (Benes, 2001). Although the maximum density of synapses may be reached as early as 15 postnatal months, it is not until 24 months that synapses develop adult morphology (Huttenlocher, 1979). There also are changes in glucose utilization and blood flow over the second half of the first year and into the second year: blood flow and glucose utilization increase above adult levels by 8e12 and 13e14 months of age, respectively (Chugani et al. Other maturational changes in prefrontal cortex, such as myelination, continue into adolescence, and adult levels of some neurotransmitters are not seen until the second and third decades of life (Benes, 2001). It is not until adolescence that neurotransmitters such as acetylcholine reach adult levels (discussed in Benes, 2001). Although much of the attention to developmental changes has focused on the medial-temporal and prefrontal regions, there also are age-related changes in the lateral temporal and parietal cortices. Cortical gray matter changes occur earlier in the frontal and occipital poles, relative to the rest of the cortex, which matures in a parietal-to-frontal direction. The superior temporal cortex is last to mature (though the temporal poles mature early; Gogtay et al. The late development of this portion of cortex is potentially significant for memory as it is one of the polymodal association areas that plays a role in integration of information across sense modalities. At a general level, we may expect concomitant behavioral development: As the neural substrate develops, so does behavior (and vice versa). But precisely how do changes in the medial temporal and cortical structures, and their interconnections, produce changes in behavior To address Memory development Chapter 18 405 this question, we must consider the basic processes involved in memory trace formation, storage, and retrieval and how the "recipe" for a memory might be affected by changes in the underlying neural substrate. In other words, we must consider how developmental changes in the substrate for memory relate to changes in the efficacy and efficiency with which information is maintained over the short term, encoded and stabilized for long-term storage, in the reliability and ease with which it is retrieved. The basic processes involved in memory are encoding, consolidation, and retrieval of memory traces. Because prefrontal cortex in particular undergoes considerable postnatal development, it is reasonable to expect that neurodevelopmental changes in it relate to age-related changes in the speed and efficiency with which information is encoded into long-term storage. Behavioral data also indicate developments in encoding throughout the second year of life. For example, relative to 15-month-olds, 12-month-olds require more trials to learn multistep events to a criterion (learning to a criterion indicates that the material was fully encoded). In turn, 15-month-olds are slower to achieve criterion, relative to 18-month-olds (Howe and Courage, 1997). Indeed, across development, older children learn more rapidly than younger children (Howe and Brainerd, 1989). For example, Rollins and Riggins (2013) observed that 6year-olds exhibited a differential pattern of neural responding during encoding, relative to adults. Furthermore, the scalp sites at which the neural responses were observed differed between children and adults, suggesting the pattern of engagement of the underlying neural substrates changes with development (Rollins and Riggins, 2013). For example, in an encoding task with children 8 years to young adulthood, Ghetti et al. The task required that participants encode both items and the colors in which they were presented. Behaviorally, whereas 8-year-olds encoded information about the items, they did not recognize the color in which the item had been presented. In contrast, 14-year-olds and college-age adults correctly recognized the items and their colors. Imaging data revealed differential patterns of activation in the hippocampus during encoding which may have contributed to the failure of the young children to "bind" the item and its color. In addition, among 10-year-olds, 14-year-olds, and adults, prefrontal activation during encoding predicted subsequent memory. These data illustrate developmental differences in encoding behavior and provide insight into the neural processes that contribute to them. In a fully mature adult with neural damage, the changes in synaptic connectivity associated with memory trace consolidation continue for hours, weeks, and even months, after an event. Memory traces are vulnerable throughout this time, as evidenced by the fact that lesions inflicted during the period of consolidation result in deficits in memory whereas lesions inflicted after a trace has been consolidated do not. For a developing organism, the period of consolidation may be one of greater vulnerability for a memory trace, relative to the adult. As a consequence, even once children have successfully encoded an event, they remain vulnerable to forgetting. Younger children may be more vulnerable to forgetting, relative to older children (Bauer, 2006a). To examine the role of consolidation and storage processes in long-term memory in 9-month-old infants, Bauer et al. After the delay, 46% of the infants evidenced ordered recall of the sequences, and 54% did not. Nevertheless, 1 week later, at the delayed recognition test, the infants who would go on to recall the events recognized the props, whereas infants who would not evidence ordered recall did not. Thus, in spite of having encoded the events, a subset of 9-month-olds failed to recognize them after 1 week and subsequently failed to recall them after 1 month. Moreover, the size of the difference in delayed recognition response predicted recall performance 1 month later. That is, infants who had stronger memory representations after a 1-week delay exhibited higher levels of recall 1 month later (see also Carver et al.
Discount entocort 200mcg on line
The phytochemical study and antibacterial activities of the seed extract of Dacryodes edulis (African Native Pear) allergy symptoms congestion generic entocort 100 mcg line. Flavonoids as anti-inflammatory agents: implications in cancer and cardiovascular disease. In vitro antioxidant activity and phytochemical screening of methanol extracts of Ficus capensis and Dacryodes edulis leaves. Tannins: current knowledge of food sources, intake, bioavailability and biological effects. Important antinutrients in plant feedstuffs for aquaculture: an update on recent findings regarding responses in salmonids. Boerhaavia diffusa inhibits key enzymes linked to type 2 diabetes in vitro and in silico; and modulates abdominal glucose absorption and muscle glucose uptake ex vivo. New antimalarial hits from Dacryodes edulis (Burseraceae)-Part I: isolation, in vitro activity, in silico "druglikeness" and pharmacokinetic profiles. Increased -cell apoptosis prevents adaptive increase in -cell mass in mouse model of type 2 diabetes: evidence for role of islet amyloid formation rather than direct action of amyloid. Dissection of metabolic, vascular, and nerve conduction interrelationships in experimental diabetic neuropathy by cyclooxygenase inhibition and acetyl-L-carnitine administration. Effect of processing methods on the antioxidant properties and inhibition of -amylase and -glucosidase by African pear (Dacryodes edulis) fruit. Altered glucose carrier expression: mechanism of intestinal adaptation during streptozocin-induced diabetes in rats. Characteristics and composition of the pulp oil and cake of the African pear, Dacryodes edulis (G. Literature re Key words: Dacryodes edulis, Traditional use, Phytochemical, Pharmacological potentialities. Epigallocatechin gallate, a constituent of green tea, represses hepatic glucose production. Induction of cancer cell apoptosis by flavonoids is associated with their ability to inhibit fatty acid synthase activity. Cytotoxicity and free radical scavenging activities of hexane fractions of Nigeria specie of African pear (Dacryodes edulis). Effect of boiling on the phytochemical constituents and antioxidant properties of African pear Dacryodes edulis seeds in vitro. Antioxidant properties of phenolic compounds: H-atom versus electron transfer mechanism. Dietary polyphenols as antioxidants and anticancer agents: more questions than answers. Protective role of epigallocatechin gallate, a dietary antioxidant against oxidative stress in various diseases the subsequent damage. Imbalance in the redox state of the cells generates toxic effects via peroxides and free radicals production. Such toxic effects damage the integral cell components such as nucleic acids, lipids, and proteins. Thiols, ascorbic acid, or polyphenol molecules are among the various antioxidants that act as reducing agents by inhibiting the oxidation of many molecules by being self-oxidized. Among these, vitamin E is the main chainbreaking antioxidant of the cell membrane. In dietary supplements, antioxidants are widely used and studies are conducted to find out the protective role of dietary supplements against various types of diseases including coronary heart disease, cancer, immunological disorders, neurodegenerative disease, and even altitude sickness. Formed by radical reactions with cellular components such as lipids and nucleobases. Oxygen-centered radicals participate in lipid peroxidation reactions produced in the presence of oxygen either by radical addition across double bonds or hydrogen abstraction. Readily oxidizes protein constituents such as thiol groups, amino groups, and methionine. It forms peroxynitrous acid upon protonation, which subsequently forms hydroxyl radical and nitrogen dioxide through homolytic cleavage. Coenzyme Q Coenzyme Q (CoQ) is naturally found in humans with high levels in the liver, heart, and kidneys, etc. CoQ is a benzoquinone derivative and is localized at the mitochondrial respiratory chain along with some other internal membranes. CoQ is directly involved in the transportation of electrons in the respiratory chain and also couples it with oxidative phosphorylation. The apoferritin shell is formed by the association of 24 ferritin subunits, where each apoferritin molecule takes up iron atoms. However, the uricase enzyme acts as an inhibitor for uric acid, but researchers have reported that such enzymes (uricase) have been inactivated during the evolution of humans and apes. Previous studies have reported that high uric acid levels in blood protect from oxidative injury in neural, vascular, and cardiac cells. Ultimately, the heme is degraded for the reduction of biliverdin to bilirubin by the biliverdinreductase enzyme. Bile pigments as potent in vitro scavengers of free radicals are an important inducible antioxidant system that protects against various cellular stresses including oxidative damage. Bilirubin protects neurons against damages caused by H2O2 and also has a protective role against ischemic injury in isolated hearts. Vitamin C is a significant antioxidant due to the formation of relatively stable ascorbyl free radical by the donation of an electron. Protective role of epigallocatechin gallate, a dietary antioxidant against oxidative stress in various diseases target molecules. Many plant pigments including -Carotene and various other flavonoids have antioxidant property12 and plant phenols are seen to inhibit lipid peroxidation and lipoxygenase enzymes (in vitro). Epigallocatechin gallate the leaves from Camellia sinensis are used in making black or green tea. Polyphenols perform the neutralization of free radicals and, hence, decrease the inflammation and inhibit the development of tumors. Study population 82 patients with early atherosclerosis 42 patients with coronary artery disease 25 male patients with wildtype transthyretin amyloid cardiomyopathy Dose 30 mL supplemented with olive oil 300 mg initial dose followed by 150 mg for 2 weeks 600 mg for 12 months Effect Improved endothelial function by reducing the number of leukocytes. Induced cytotoxicity of human sebocytes via apoptosis and decreased the viability of P. Epigallocatechin gallate and cancer Oncogenesis is considered as a composite and several-step procedure where modifications take place at the cellular and molecular levels. Protective role of epigallocatechin gallate, a dietary antioxidant against oxidative stress in various diseases progression stage, known as the neoplastic transformation stage where uncontrolled growth leads to tumor formation. This stage involves the formation of an abnormal mass of tissue (neoplastic) from the premalignant cells with improved potential of metastasis, invasiveness, and angiogenesis. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Uric acid provides an antioxidant defense in humans against oxidant- and radical-caused aging and cancer: a hypothesis. Heme oxygenase-1-derived bilirubin ameliorates postischemic myocardial dysfunction. Green tea and its polyphenolic catechins: medicinal uses in cancer and noncancer applications. Epigallocatechin, a green tea polyphenol, attenuates myocardial ischemia reperfusion injury in rats. Green tea epigallocatechin-3-gallate attenuates Porphyromonas gingivalisinduced atherosclerosis. Epigallocatechin-3-gallate prevents cardiac apoptosis by modulating the intrinsic apoptotic pathway in isoproterenol-induced myocardial infarction. Polyphenol (-)-epigallocatechin gallate-induced cardioprotection may attenuate ischemia-reperfusion injury through adenosine receptor activation: a preliminary study. Beneficial effects of polyphenol-rich olive oil in patients with early atherosclerosis.
Cheap 200 mcg entocort overnight delivery
Sequence of theory-of-mind acquisition in Turkish children from diverse social backgrounds allergy forecast new jersey generic 100 mcg entocort visa. Preverbal infants identify emotional reactions that are incongruent with goal outcomes. Neural evidence that three dimensions organize mental state representation: rationality, social impact, and valence. Inscapes: a movie paradigm to improve compliance in functional magnetic resonance imaging. Beyond superior temporal cortex: intersubject correlations in narrative speech comprehension. Oxytocin receptor gene relates to theory of mind and prosocial behavior in children. One-to four-year-olds connect diverse positive emotional vocalizations to their probable causes. Disruption of the right temporoparietal junction with transcranial magnetic stimulation reduces the role of beliefs in moral judgments. Chapter 22 A developmental neuroscience perspective on empathy Jean Decety1, 2 and Kalina J. Conclusions List of abbreviations References 493 494 494 496 496 498 499 499 500 22. Among the psychological processes that are the basis for much of sociability and smooth social interaction, empathy plays a pivotal role. Empathy-related responding, including caring and sympathetic concern, motivates bonding between individuals and some forms of prosocial behavior, inhibits aggression, and overall facilitates group living and cooperation. On the other hand, certain developmental disorders are marked by empathy deficits, which influence the motivation to respond to others in distress or need, and care for them. Understanding how these motivations and behaviors are implemented in the brain and downstream peripheral physiology, both in typically developing children and children with antisocial tendencies, can help elucidate the role of empathy in prosociality. It is important to note that empathy can compete with moral judgment and justice principles, for instance, by inducing partiality for in-group members (Cikara and Van Bavel, 2014; Decety and Cowell, 2014a). This article critically examines the current knowledge about the development of the mechanisms supporting empathy and its associated behavioral responses such as some forms of prosocial behaviors including caring. We review the affective and cognitive components that give rise to empathy, starting first with the automatic proclivity to share affect and emotions with others, and then the cognitive processes of perspective-taking and executive control, which enable individuals to intentionally adopt the subjective view point of another without confusion between self and other. Based on conceptual and empirical evidence from developmental psychology, cognitive neuroscience, and neurology, a number of distinct and interacting components contribute to the experience of empathy: (1) affect sharing, a bottom-up process grounded in affective arousal and neural circuits connecting the brain stem, amygdala, basal ganglia, and orbitofrontal cortex, (2) understanding emotion that relies on self- and other-awareness and critically involves the medial and ventromedial prefrontal cortex and temporoparietal junction, and (3) executive functions instantiated in the prefrontal cortex that operate as a top-down mediator, allowing for perspective-taking; emotion regulation, and appraisal of social context (Decety and Jackson, 2004; Decety and Meyer, 2008). Drawing from multiple sources of data and levels of analysis provides a more complete picture of the phenomenological experience of empathy, as well as an understanding of the development and interaction between the mechanisms that drive the phenomenon. Furthermore, studying subcomponents of more complex psychological constructs such as empathy can be particularly useful from a developmental perspective, because only some of its components or precursors may be observable. Developmental studies can provide unique opportunities to see how the components of the system interact in ways that are not always possible in adultsdwhere all the components are fully mature and operational. Until quite recently, research on the development of empathy-related responding from a neurobiological level of analysis has been relatively sparse. We believe that integrating this perspective with behavioral observations can shed light into the neurobiological mechanisms underpinning the basic building blocks of empathy and sympathy and their age-related functional changes. Such integration helps us characterize the neurobiological processes that underpin interpersonal affective responding and prosocial behavior while also potentially informing interventions for individuals with atypical development, such as antisocial behavior problems. Empathy is implemented by a network of distributed, recursively connected, interacting neural systems and regions, including the superior temporal sulcus, insula, medial and orbitofrontal cortices, amygdala, and anterior cingulate cortex, as well as autonomic and neuroendocrine systems implicated in sociality. Prosocial behavior is an umbrella term for actions that are benefiting another person. This umbrella concept includes many different types of behaviors such as helping, cooperating, sharing, comforting, rescuing, and informing. These various forms of prosocial behaviors have distinct underlying motivations such as caring, fairness, reputation management, group loyalty, reciprocity, social rewards, etc. Morality refers to prescriptive norms regarding how people should treat one another, including concepts such as justice, fairness, and rights. All definitions of morality minimally include judgment of the rightness or wrongness of acts or behaviors that knowingly cause harm to people. Autonomic synchronization involves any associative pattern in the physiologies of interacting partner like a mother and her child, such as synchrony in heart rate, respiration rhythm, pupil diameter, and hormonal level. Emotion contagion is the unconscious tendency to take on the sensory, motor, physiological, and affective states of others. Neurodevelopmental studies provide unique opportunities to explore how the components of empathic responding interact in ways that are not possible in adults. Investigating dysfunction of the components of empathy provides important clues for understanding deviations that can lead to the lack of concern for others in social decision-making and behavior. Some scholars more narrowly characterize empathy as one specific set of congruent A developmental neuroscience perspective on empathy Chapter 22 487 emotions, those feelings that are more other-focused than self-focused, and employ the notion of empathic concern (Batson, 2012), which is functionally linked with a motivation to care for the welfare of another. The experience of empathy can lead to caring (an other-oriented motivation) or personal distress (an egoistic motivation to reduce stress by withdrawing from the stressor, thereby decreasing the likelihood of prosocial behavior). Emotion regulation is considered to be a critical component of empathy, as the modulation of emotional experience allows an individual to remain aware of an emotionally evocative situation without being overwhelmed or numbed by it. This is particularly important in the case of negative arousal (Decety and Lamm, 2009). Developmental research indicates that children who are able to regulate their emotions are also more likely to experience sympathy (concern) rather than personal distress (Eisenberg and Eggum, 2009). For instance, in 5- to 6-year-old children, prosocial behavior is significantly correlated with ratings of the emotional state of the protagonist but not with own emotional state, suggesting that empathic concern rather than personal distress is the primary influence on prosocial behavior (Williams et al. These facets operate by way of automatic and controlled processes and interact with one another. Yet they can be dissociated, as they rely on partially separable information processing systems and underlie different functions (Shdo et al. Moreover, these facets have different developmental trajectories and phylogenetic roots (Decety and Svetlova, 2012). In particular, there is evidence for agerelated changes in these neural circuits, which, together with behavioral observations, reflect how brain maturation influences the reaction to the distress of others (Decety and Michalska, 2010; Levy et al. Functional imaging studies reveal that the anterior insula and the anterior cingulate cortices are conjointly activated during the experience of negative emotion and during the perception of negative emotion in others. The insula provides a foundation for the representation of subjective bodily feelings, which substantiates emotional awareness. For instance, current mood states, relationship to the person, social status, and the context in which the interaction occurs influence the manner and the extent to which the observer will react. In the following sections, each of the components of empathy (affect sharing, emotion understanding, perspective-taking, and emotion regulation) will be considered separately from a developmental neuroscience perspective. These components are dissociable as documented in patients with brain lesions (Shdo et al. In addition, both genetic and environmental factors contribute to the development of empathy and prosociality (Knafo et al. Both developmentally and evolutionarily, advanced forms of empathy are preceded by and grow out of more elementary ones, such as the capacity to express and respond adaptively to emotional signals (Decety et al. Affective responsiveness is known to be present at an early age, is involuntary, and relies on somatosensorimotor resonance between other and self (Decety and Meyer, 2008). This type of physiological linkage is shared by most mammals and represents the earliest form of emotional contagion that occurs between a mother and a child even before birth (Feldman, 2016). Infants implicitly pick up these subtle social signals from their caregivers, and this in turn has an impact on their own physiology and cognition (Prochazkova and Kret, 2017). The experience of synchrony during the first months of age demonstrates how critical environmental input is for both the maturation of neural circuits that support social engagement (Johnson et al. In a cross-sectional study, 6- and 9-month-old infants were presented with schematic depictions of eyes with smaller and larger pupils while their own pupil sizes were recorded (Fawcett et al. Similarly, another study demonstrated that 6- and 12-month-old infants had greater pupil dilation while viewing videos of other infants laughing and crying than while viewing videos of neutral babbling (Geangu et al. This spontaneous transfer of internal states is fundamental for survival, social group cohesion and, we contend, the development of empathy.
Order entocort 200mcg without prescription
Some types of memory have a limited capacity allergy testing for gluten cheap entocort 200mcg free shipping, whereas others are for all practical purposes, limitless. In fact, it sometimes seems that the only thing all memories have in common is that it is about the past. Yet even this characterization is not universally true, in that remembering to do something in the future also qualifies as "memory," of the prospective type. This step is necessary in order to accomplish the second goal, which is to describe the course of development of different types of memory over the first two decades of life. Because it features some of the most profound changes, the focus will be on long-term memory. As will become apparent, different types of long-term memory have different courses of development. Differences in the timing and course of development shed light on some of the mechanisms of age-related changes in childhood, consideration of which is the third major goal of the chapter. Maine de Biran (1804/1929) is credited as the first to suggest that there might be different forms of memory (Schacter et al. The notion was furthered at the beginning of the 20th century with studies of wounded veterans from World War I. Kleist (1934), a German physician, examined veterans who had received head wounds from gun shots or shrapnel and the behavioral patterns that seemed to result from them. He observed that there were systematic relations between the site of the wound (and resulting brain lesion) and the 395 Neural Circuit and Cognitive Development. The notion that different parts of the brain subserve different cognitive functions, including different types of memory, received especially strong impetus from the famous case of H. To treat the seizures, his surgeon removed large portions of the temporal lobes on both sides of the brain (Scoville and Milner, 1957). His case in particular led to the notion that there are different types or forms of memory. Whereas some distinctions within the domain are readily accepted, others are sources of debate. Whereas some memories are short term, lasting only seconds, others last much longer, on the order of days, months, years, and perhaps even a lifetime. Historically it was thought to hold 7 "units" of informationdsuch as digits in a phone numberdplus or minus 2 (Miller, 1956). More recent conceptualizations of short-term memory purport a store of only 3e5 units, depending on the contents of each unit and how they are processed. There seemingly is no ceiling on the number of items, pieces of information, or personal experiences, that one can maintain in long-term memory stores (more on the temporal dimension and capacity limitations of memory below). Although the precise distinctions captured by the different labels given to the divisions are not identical, there is substantial overlap in contemporary conceptualizations of declarative or explicit memory versus nondeclarative, procedural, or implicit memory. These are entities that we think of as being encoded symbolically and that thus can be described with language. In terms of function, declarative memory is specialized for fast processing and learning. New information can be entered into the declarative memory system on the basis of a single trial or experience. In terms of rules of operation, declarative memory is fallible: we forget names, dates, and places, for example. Although there are compelling demonstrations of long-term remembering of lessons learned in high school and college. Memory development Chapter 18 397 subsequent retrieval of declarative or explicit memories depends on a multicomponent network involving cortical structures as well as medial temporal structures. Declarative memory is itself subdivided into the categories of semantic memory and episodic memory. We are consulting semantic memory when we retrieve the facts that the Appalachian Trail is 2190 miles long and crosses 14 states, the southern terminus of the Appalachian Trail is in Georgia, and that the idea for the trail was developed by Benton MacKaye and proposed in 1921. For practical purposes, both the capacity of semantic memory and the longevity of the information stored in it seem infinite. That is, we know facts and figures, names and dates, yet in most cases, we do not know when and where we learned this information. We might be able to reconstruct how old we were or what grade we were in when we learned some tidbits of information, but unless there was something unique about the experience surrounding the acquisition of this information, we carry it around without address or reference to a specific episode. In contrast to semantic memory, episodic memory supports retention of information about unique events (Tulving, 1972, 1983), such as hiking a section of the Appalachian Trail or the fact that Georgia was one of the states on a list of state names studied in a memory experiment. Some episodic memories, such as whether a specific state was included in a word list, may not stay with us for very long, and are not especially personally relevant or significant. These so-called autobiographical memories are episodic memories that are infused with a sense of personal involvement or ownership (Bauer, 2007, 2015). They are the episodes on which we reflect when we consider who we are and how our previous experiences have shaped us. Skilled motor behavior, such as playing piano or jumping hurdles, is not a name, date, place, fact, or event, but a collection of finely tuned motor patterns, behaviors, and perceptual skills that we cannot verbally describe. Most types of nondeclarative memory function to support gradual, incremental learning. Perhaps as a result of its slow function, a rule of the operation of nondeclarative memory is that the learning is relatively infallible. A typical example is riding a bicycledwe may not have ridden a bicycle in many years but when we ride one again, we "just know how. As might be expected, given the diversity of behaviors supported by nondeclarative memory, different types of nondeclarative memory depend on different neural structures. For example, motor skill learning and many types of conditioning seemingly are dependent on cerebellum and subcortical structures such as the basal ganglia, and priming seems dependent on extrastriate cortex. The most common categories of nondeclarative memory are (a) motor skill learning, (b) priming, and (c) classical conditioning. What they have in common is that although there might be a declarative component to them (to stop the bike, put on the brakes), their fluid execution is not accomplished via learning and frequent repetition of the declarative "rules," but motor practice. Behind an exquisite piano performance or a championship winning jump are innumerable hours of practicedrepetition after repetition of a passage of music or agility drills. One can learn a simple tune like "Mary Had a Little Lamb" or basic jumping movements in an elementary class, but it is not this knowledge on which experts depend. Potatoes, onions, celery, and carrots must all be chopped before going into the pot, but each requires a different amount of pressure to slice through successfully. Luckily, we do not need to consciously consult our memory to apply the appropriate pressure each and every time. The information is encoded in our muscles and joints; it is not accessible to conscious access or description. Another form of nondeclarative memory is priming, which is facilitated processing of a stimulus following prior exposure to the stimulus. Perceptual priming occurs when subsequent processing of a stimulus is facilitated by prior perceptual exposure to it. Conceptual priming occurs when there is overlap between related concepts in memory. A person who had recently studied a list that included the word "Alaska" would be more likely to include it in a list of states than an individual who had not studied the item. With regard to the distinction between conscious (declarative) and unconscious (nondeclarative) memory, an important point to remember about priming is that it can occur without conscious awareness that the item had been studied earlier, or that subsequent processing was facilitated by earlier processing. That is, facilitated processing occurs even in the absence of recognition or recall of the originally studied item (see Lloyd and Newcombe, 2009; Roediger and McDermott, 1993, for discussions). Typically, one of the stimulidthe unconditioned stimulusdevokes a high-probability or even reflexive response termed the unconditioned response. For example, a puff of air to the eye (unconditioned stimulus) produces a blink (unconditioned response). The other stimulusdthe conditioned stimulusdis behaviorally neutral, such as a tone. Classical conditioning occurs when, over time and repeated pairing of the two stimuli, the conditioned stimulus (the tone) takes on the same significance as the unconditioned stimulus, such that it alone is sufficient to produce the response: anticipatory eye closure at the sound of the tone (see Woodruff-Pak and Disterhoft, 2008, for a review). Although we may be consciously aware that vehicle stopping distance is 15 feet per second, it is not this knowledge that permits us to bring our vehicles to a gentle stop at a stop sign.
