

Order viagra sublingual 100mg overnight delivery
In other words erectile dysfunction medication samples buy viagra sublingual 100 mg without prescription, the output is highly sensitive to changes in the level of ComK (the opposite of robustness). Whether or not cells have the potential to turn on comK is governed by a regulatory pathway operating at the level of the proteolytic stability of the ComK protein. That is, under conditions in which ComK is not subject to degradation, only some of the cells in the population become competent. This can be vividly seen using cells harboring a fluorescent reporter (the gene for the green fluorescent protein) for ComK-directed gene activity. Thus, noise in the expression of the comK gene resulting in small variations in the levels of ComK between cells enables the activator to reach a threshold concentration in some cells and not others. This example of positive autoregulation illustrates how noise in gene expression can be exploited to drive cells in to alternative states. Returning to the language of systems biology, we would say that bacteriophage l has a two-node switch linked in both directions by negative edges. Although numerous examples of bistable switches are found in bacteria, bistability is by no means limited to microbes. A double-negative-feedback loop that can be stably maintained in one state or another dictates whether a common precursor cell will express one set of receptors or the other. Rather, upstream signals dictate in which direction the switch is thrown, whereas the double-negativefeedback loop subsequently locks the switch in its predetermined state. Bimodal Switches Vary in Their Persistence Bistable switches, as we have seen, are bimodal in that they can persist for extended periods of time in alternative stable states. Some regulatory circuits that show bimodality are said to be excitable because they do not persist in alternative stable states. Like bistable systems, excitable systems involve a self-reinforcing circuit that causes a large and stereotypical response to a small perturbation. In cells harboring the modified gene alone, no bistability is observed, and the level of ComK-directed gene expression increases in a more or less uniform manner in response to increasing levels of inducer, showing a unimodal distribution of expression levels among cells in the population at any give concentration of inducer (Box 22-1. However, in cells harboring both the modified gene and the normal autoregulated gene, increasing concentrations of inducer cause the cells to bifurcate in to a subpopulation showing a low level of ComK activity and a subpopulation showing a high level of ComK activity (Box 22-1. Strictly speaking, the use of the term "bistability" requires that a switch show a property called hysteresis. When exposed to a magnetic field, the material becomes magnetized and, importantly, remains so even when the external magnetic field is removed. Now let us return to our example of cells harboring both ComK and a modified copy of ComK that responds to an inducer. Now consider what happens when we lower the level of inducer such that less and less ComK is produced from the engineered copy of the gene. The switch governing the decision between the lysogenic and lytic modes of propagation of bacteriophage l is also hysteretic. Note that the cells in a and b harbored a fusion of the gene for the green fluorescent protein to a promoter under the control of ComK. A classic example of an excitable system in biology is the action potential of a neuron. This inward flow of positive ions causes the membrane potential to rise still higher, triggering additional channels that had not yet opened to allow sodium ions in to the cell. Eventually, this cascade of channel openings culminates in a peak of positive voltage (40 mV) inside the cell. Excess sodium ions are then pumped out of the neuron, restoring the membrane to its original resting state. Thus, a small perturbation in membrane potential triggers a large, programmed response but one that sets in motion its own rapid reversal to the original resting state. Likewise, self-reinforcing regulatory circuits that are unable to sustain alternative states for extended periods of time or that set in motion a chain of events that causes the circuit to reverse can be considered to be excitable. Positive autoregulation by ComK creates a bistable switch that could maintain ComK at high levels for an extended period of time. However, superimposed on self-reinforcing synthesis of ComK is a negative-feedback loop that eventually leads to the proteolytic destruction of the activator protein. This negative-feedback circuit enables competent cells to exit their non-growing, competence state and return to a proliferative, vegetative state. The phage l genetic switch, in contrast, can maintain the lysogenic state for many generations and hence is most properly considered to be bistable. A much more robust example of excitability is shown by the very same bacterium when it is actively growing. Importantly, the two cell types switch back and forth stochastically at a frequency of tens of cell generations. One hypothesis is that the bacterium has evolved to hedge its bets, not knowing how long current favorable conditions might last. The sessile chains can be thought of as settlers that stick to surfaces and exploit a currently favorable microenvironment, whereas the swimmers are foragers that swim off in search of new, favorable environments. For simplicity, SlrR and SinR are abbreviated to Slr and R, respectively, in the cartoon. The kymograph on the right shows a time-lapse series of micrographs taken at 5-min intervals. A motile cell (green) at the bottom left switches to the chaining state (red), giving rise to progeny that maintain the chaining state. Eventually, a chaining cell near the bottom switches back to the motility state (green), giving rise to progeny that maintain the motility state. This switching can be visualized in real time using a microfluidic device in which cells are embedded in long channels, each the width of a bacterium. The cells carry a fluorescent reporter for a motility gene (green) and for a matrix gene (red) that is characteristic of the chaining state. A motile (green) cell at the bottom of the channel gave rise to progeny that expressed chaining genes (red). Subsequently, a cell in the chaining state near the bottom switches back to a cell expressing motility genes (green). Data collection from large numbers of time-lapse experiments show that cells persist for multiple generations in each state and switch from one to the other stochastically. Evidently, certain circuits have beneficial properties that are favored by natural selection. Feed-Forward Loops Are Three-Node Networks with Beneficial Properties A striking example of this is provided by networks that consist of three nodes. These can be distinguished from each other by the direction of the edges, whether edges connect two or all three nodes, and whether pairs of nodes are connected by one or two edges. We refer to it as a "network motif" because it is a recurring theme in genetic circuits. Again, natural selection has favored two that are found more commonly than the others. In one of the favored feed-forward loop motifs (known as a "coherent motif"), both the direct and the indirect pathways leading to the target gene, representing the output, have the same sign. In the other favored motif (known as an incoherent motif), the two pathways have different signs, with the target gene C being subject to positive control by A in the direct pathway and negative control by B in the indirect pathway. Because both motifs are favored among all other feed-forward loops and indeed among all possible three-node networks, it is reasonable to expect that they have favorable properties that have been the basis for their selection in evolution. Indeed, computational modeling and experiment reveal that each motif has characteristics that make them useful in regulatory circuits. For example, the coherent feed-forward loop has the property of requiring a sustained input in order for the target gene C to be transcribed. In other words, this kind of feed-forward loop is a persistence detector that only responds to a signal that is long-lived or persistent. This property derives from the fact that turning on the target gene depends on both the primary activator A and sufficient accumulation of the secondary activator B. Thus, the input signal must persist long enough for the secondary activator B to reach the threshold concentration needed to turn on the target gene C.
Syndromes
- Discomfort during the test
- Cold sweat
- Blood clots in the lung (pulmonary embolism)
- Fluid buildup in the knee joint
- Over-filling the bladder with fluid while under general anesthesia, called bladder hydrodistention
- Weakness
- Fever
- Antinuclear antibody panel (ANA)
- Limiting alcohol
- Chest x-ray
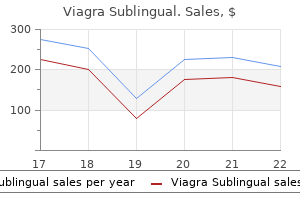
100 mg viagra sublingual otc
This is how we know that the proteins are not needed for the splicing reaction itself impotence 101 purchase viagra sublingual 100 mg free shipping. The similar chemistry seen in self- and spliceosome-mediated splicing is believed to reflect an evolutionary relationship. The proposed striking similarities in these structures have since been confirmed through X-ray crystallographic studies. In this way, introns had only to retain the minimum of sequence elements required to target splicing to the correct places. In the case of the spliceosome and the ribosome, however, these activities have not been entirely replaced by proteins. This happens if the spliceosome components bound at the 50 splice site of one exon interact with spliceosome components bound at the 30 splice site of not the next exon, but one beyond. In the case shown, the pseudo-site is within an exon and leads to regions near the 50 end of that exon being mistakenly spliced out along with the intron. It is unlikely both would recognize an incorrect sequence, and thus selection is stringent. Consider the following: the average human gene has seven or eight exons and can be spliced in three alternative forms. But there is one human gene with 363 exons and one Drosophila gene that can be spliced in 38,000 alternative ways, a case we describe in detail in the next section. Remember also that the average exon is only some 150 nucleotides long, whereas the average intron is 3000 nucleotides long (and as we have seen, some introns can be as long as 800,000 nucleotides). First, splice sites can be skipped, with components bound at, for example, a given 50 splice site pairing with those at a 30 site beyond the correct one. Second, other sites, close in sequence but not legitimate splice sites, could be mistakenly recognized. This is easy to appreciate when one recalls that the splice site consensus sequences are rather loose. Therefore, for example, components at a given 50 splice site might pair with components bound incorrectly at such a "pseudo" 30 splice site. Once in place, the 50 splice site components are poised to interact with those other factors that bind to the next 30 splice site to be synthesized. Thus, the correct 30 splice site can be recognized before any competing sites further downstream have been transcribed. This cotranscriptional loading process greatly diminishes the likelihood of exon skipping. Legitimate splice sites are recognized by the splicing machinery by virtue of being close to exons. This initiates the assembly of the splicing machinery on the correct sites, and splicing can proceed as outlined above. In this way, the machinery binds more efficiently to those nearby splice sites than to incorrect sites not close to exons. As we saw above, these factors demarcate the splice sites for the rest of the machinery to assemble correctly. By recruiting splicing factors to each side of a given exon, this process encourages the so-called "exon definition" we alluded to above when discussing the order of events during spliceosome assembly. That is, spliceosome components are recruited around exons initially, rather than around the intron to be removed. Subsequently, components near one exon will pair with those near an adjacent exon to eliminate the intervening intron. They not only ensure the accuracy and efficiency of constitutive splicing (as we have just seen) but also regulate alternative splicing (as we shall see presently). They come in many varieties, some controlled by physiological signals, others constitutively active. Some are expressed preferentially in certain cell types and control splicing in cell-type-specific patterns. The chemistry of this reaction is the same as that of the standard splicing reaction described above, and the spliced product is indistinguishable. The only difference is that the other product- the lariat in the standard reaction-is, in trans-splicing, a Y-shaped branch structure instead. In alternative splicing, exons can be deliberately skipped, and a given exon is joined to one further downstream (as we shall see later). Trans-splicing uses the same spliceosomal machinery as normal cis-splicing, except for U1, which, at least in worms, is not needed for trans-splicing. This rare form contains some components common to the major spliceosome, but it contains other unique components as well. Thus, U11 and U12 components of the alternative spliceosome have the same roles in the splicing reaction as U1 and U2 of the major form, but they recognize distinct sequences. Finally, the identical U5 component is found in both the major and the alternative-so-called minor-spliceosome. It should be emphasized that although these introns are rare, they are widely distributed-approximately 800 human genes contain at least one minor intron. Despite the different splice site and branch site sequences recognized by the two systems, these major and minor forms of spliceosomes both remove introns using the same chemical pathway. Regardless, the chemistry is the same, and so are some of the spliceosome components, and others are closely related. It is now believed that at least 40% of Drosophila genes and as many as 90% of human genes undergo alternative splicing. Shown is a region of the troponin T gene encoding five exons that generates two alternatively spliced forms as indicated. Alternative splicing is sometimes used as a way of generating diversity, with alternative forms being generated stochastically. But in many cases, the process is regulated to ensure that different protein products are made in different cell types or in response to different conditions. For a simple case of alternative splicing, consider the gene for the mammalian muscle protein troponin T. In other cases, exons can be skipped (deliberately), or introns can be retained in the mature message. Some alternative spicing results from transcription of a gene from alternative promoters, allowing one transcript to include a 50 exon not present in the other. Similarly, alternative poly-A sites allow 30 terminal exons to be extended or alternative 30 terminal exons to be used in some transcripts of a given gene. The T-antigen gene encodes two protein products: the large T antigen (T-ag) and the small t antigen (t-ag). Large T induces transformation and cell cycle reentry, whereas small t blocks the apoptotic response of cells forced down that path. In genome-wide studies, the most commonly seen forms of alternative splicing are cases in which complete exons are included or excluded from the mature message. In these cases, there must be mechanisms that ensure that the exons are spliced in a mutually exclusive fashion. Several Mechanisms Exist to Ensure Mutually Exclusive Splicing There are several mechanisms to ensure that selection of alternative exons is mutually exclusive-that is, that when one is chosen, the other is not (or, to refer again to the a-troponin T example, when exon 3 is chosen, exon 4 is always excluded, and vice versa). We deal with each of these mechanisms here and then, in the next section, discuss an extreme case in which a special mechanism is required. If the splice sites within the intron are too close together, splicing factors cannot bind to both sites at the same time. The splicing of exons 3 and 4 of a-troponin is made mutually exclusive by this mechanism. This arrangement can arise through the relative positions of the splice sites within an intron or because the intron is simply too small to work; in Drosophila, any intron under 59 nucleotides falls in to that category. Combinations of Major and Minor Splice Sites As we saw above, there is a form of the spliceosome called the minor spliceosome that recognizes splice sites distinct from those recognized by the major spliceosome. Nonsense-Mediated Decay Rather than forcing the splicing machinery to splice in a mutually exclusive fashion, this mechanism instead ensures that only messages that have one or another exon (never both and never neither) survive. In other words, although not ensuring mutually exclusive splicing, the consequences of this mechanism amount to the same thing. In this case, binding of U1 to the 50 splice site of the first intron results in exclusion of exon 2. The Curious Case of the Drosophila Dscam Gene: Mutually Exclusive Splicing on a Grand Scale the Drosophila Dscam (Down syndrome cell-adhesion molecule) gene potentially encodes 38,016 protein isoforms.
Generic viagra sublingual 100 mg otc
Then impotence urology cheap viagra sublingual 100 mg with mastercard, in preparation for the first nuclear division, these duplicated homologous chromosomes must pair and align at the center of the cell. Recombination must be complete before the first nuclear division to allow the homologs to properly align and then separate. The products of this division are the four gametes, each with one copy of each chromosome. In the absence of recombination, chromosomes often fail to align properly for the first meiotic division, and, as a result, there is a high incidence of chromosome loss. This improper segregation of chromosomes, called nondisjunction, leads to a large number of gametes without the correct chromosome complement. Gametes with either too few or too many chromosomes cannot develop properly once fertilized; thus, a failure in homologous recombination is often reflected in poor fertility. The homologous recombination events that occur during meiosis are called meiotic recombination. Meiotic recombination also frequently gives rise to crossing over between genes on the two homologous parental chromosomes. The pairs are called sister chromatids, and recombination that occurs between non-sister pairs is mediated by Dmc1. Rather, the cut sites are located most commonly in chromosomal regions that are not tightly packed with nucleosomes, such as promoters controlling gene transcription (see Chapters 8 and 19). After two generations of growth in bromodeoxyuridine, one newly replicated chromatid has only one of its strands substituted, whereas its sister has both substituted. Then, crossovers are easily detected as alternating lengths of light and dark (a). Protein sequence comparisons reveal that Spo11 appears to be a distant cousin of these enzymes. This resealing can occur when cells receive a signal to stop proceeding with meiosis. Both proteins participate in recombination, but how they work together is not known. Whereas Rad51 is widely expressed in cells dividing mitotically and meiotically, Dmc1 is expressed only as cells enter meiosis. Interestingly, Dmc1dependent recombination is preferentially between the non-sister homologous chromatids, rather than between the sisters. Although the mechanistic basis of this selectivity is unknown, there is a clear biological rationale: Meiotic recombination promotes interhomolog connections to assist alignment of the chromosomes for division. Genetic experiments indicate that many additional proteins also participate in this process. In addition, many proteins appear to interact with the known recombination enzymes, and it seems likely that these proteins function in the context of a large multicomponent complex. Proteins were detected by immunostaining with fluorescently labeled antibodies to Rad51 (green) and Dmc1 (red). By analogy with bacteria, we expect that eukaryotic cells encode proteins that promote the branch migration and Holliday junction resolution steps of recombination. In fact, enzymes capable of promoting these reactions are being identified and characterized. In humans, for example, an alternative process to resolve a double Holliday junction involves a RecQ helicase acting in concert with a topoisomerase. This mechanism, called double-junction dissolution, prevents the exchange of flanking sequences. As we have seen, meiotic recombination aligns homologous chromosomes and promotes genetic exchange between them. Specifically, these helicases can process and edit recombination intermediates, often resulting in the collapse of joint molecules before establishment of the double Holliday junction intermediate. As a result, the helicases promote non- crossover recombination at the expensive of the crossover class of events (Box 11-3. In the crossover pathway (upper pathway shown here), the Holliday junction resolvase (shown as scissors) assembles at the junction and cleaves asymmetrically to produce the crossover products. In contrast, in the non-crossover pathway (lower pathway), the RecQ-family helicase (noted by green arrow) promotes synthesis-dependent strand annealing and resolution. The mechanism of action appears to be that the helicase activity of these enzymes takes apart the joint molecules initially made by the strand exchange proteins. These events may provide the essential chromosome-pairing function needed for a successful meiotic division, yet leave no detectable change in the genetic makeup of the chromosomes. But even noncrossover recombination can have genetic consequences, such as giving rise to a gene conversion event. Gene conversion happens Homologous Recombination at the Molecular Level 369 when an allele of a gene is lost and replaced by an alternative allele. Examples of how gene conversion can occur both in mitotically growing cells and during meiosis are described in the following sections. In addition, when an a cell and an a cell come in to close proximity, they can fuse. The a/a cell may then go through meiosis to form two haploid a cells and two haploid a cells. These regulators control expression of target genes whose products define each cell type. The regulators encoded by the mating-type genes, together with others found in all three cell types, act in various combinations to ensure that the correct pattern of genes is expressed in each cell type (see Chapter 19). In contrast, the strands terminating with 30 ends are very stable and not subjected to nuclease digestion. This type of nonreciprocal recombination event is a specialized example of gene conversion. One hint that the mechanism is distinct is that the crossover class of recombination products is never observed Homologous Recombination at the Molecular Level 371 during mating-type switching. Therefore, models for recombination that do not involve resolution of Holliday junction intermediates better explain mating-type switching. Once this annealing step occurs, the corresponding long 30 tail (bottom strand in. This mechanism nicely explains how gene conversion occurs without the need to cleave a Holliday junction. With this model, the absence of crossover products during mating-type recombination is no longer a mystery. This fundamental aspect of homologous recombination is what makes it possible to use recombination frequencies to generate useful genetic maps that display the order and spacing of genes along a chromosome. Regions with a higher than average probability are "hot spots," whereas regions that participate less commonly than an average segment are "cold. In contrast, genes separated by a "cold" interval appear by genetic mapping to be closer together than is true from their physical distance. We have encountered two examples for the molecular explanation of hot and cold spots in chromosomes. Regions near Chi sites and Spo11 cleavage sites have a higher than average probability of initiating recombination and are "hot," whereas regions having few such sites are correspondingly "cold. We have introduced the concept of gene conversion during the specialized recombination events responsible for mating-type switching in yeast. However, gene conversion is also commonly observed during normal homologous recombination events, such as those responsible for genetic exchange in bacteria and for pairing chromosomes during meiosis. To illustrate gene conversion during meiotic recombination, consider a cell undergoing meiosis that has the A allele on one homolog and the a allele on the other. In the absence of gene conversion, two gametes carrying the A allele and two gametes carrying the a allele would be generated. Notice in the region between Spo7 and Cdc15 that the genetic map is contracted because of a low frequency of crossing over. The second mechanism of gene conversion involves the repair of basepair mismatches that occur in the recombination intermediates. When working on recombination intermediates, the mismatch repair enzymes will likely choose randomly which strand to repair. Therefore, after their action, both strands will carry the sequence encoding either the A information or the a information (depending on which strand was "fixed" by the repair enzymes), and gene conversion will be observed. The double-strand repair pathway of homologous recombination well describes many recombination events. Additional enzymes promote the movement (branch migration) and cleavage (resolution) of Holliday junctions.
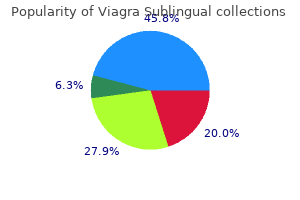
Buy viagra sublingual master card
In this case erectile dysfunction treatment photos generic 100 mg viagra sublingual with visa, the bromodomain facilitates the maintenance and propagation of acetylated histones by modifying nucleosomes that are adjacent to the already acetylated histones (as we shall discuss later). Many nucleosome-remodeling complexes include one or more subunits with domains that recognize modified histones (see Table 8-6) allowing modified histones to recruit these enzymes. This property can be used to map nucleosomes that are associated with the same position throughout a cell population (Box 8-3. To map nucleosome location accurately, it is important to isolate the cellular chromatin and treat it with the appropriate amount of micrococcal nuclease with minimal disruption of the overall chromatin structure. This is typically achieved by gently lysing cells while leaving the nuclei intact. The nuclei are then briefly treated (typically for 1 min) with several different concentrations of micrococcal nuclease, a protein small enough to diffuse rapidly in to the nucleus. The goal of the titration is for micrococcal nuclease to cleave the region of interest only once in each cell. More recently, a related approach has been developed to identify positioned nucleosomes across entire genomes. Using this approach, all of the positioned nucleosomes across an entire genome can be mapped. The experimental steps in determining nucleosome positioning in the cell are illustrated. Specific Enzymes Are Responsible for Histone Modification the histone modifications we have just described are dynamic and are catalyzed by specific enzymes. A number of different histone acetyltransferases and deacetylases have been identified and are distinguished by their abilities Genome Structure, Chromatin, and the Nucleosome 249 to target a different subset of histones or in some cases specific lysines in one histone. Histone methyltransferases and demethylases appear to be much more specific, always targeting only one of the many lysines or arginines on a specific histone (Table 8-7). Like their nucleosome-remodeling complex counterparts, these modifying enzymes are part of large multiprotein complexes. As discussed in Chapters 13 and 19, the protein complexes involved in these modifications are frequently recruited to sites of active transcription. Modification of amino-terminal tails can reduce the ability of nucleosome arrays to form repressive structures. This change creates sites that can recruit other proteins, including nucleosome remodelers. As discussed above, the first step in the assembly of a nucleosome is the binding of an H3. Although we show the order of association as histone acetylation complex and then nucleosome-remodeling complex, both orders are observed and can be equally effective. It is also true that recruitment of a different histone-modifying complex could result in the formation of more compact and inaccessible chromatin. To duplicate a chromosome, at least half of the nucleosomes on the daughter chromosomes must be newly synthesized. Are all of the old histones lost and only new histones assembled in to nucleosomes If not, how are the old histones distributed between the two daughter chromosomes The fate of the old histones is a particularly important issue given the effects that histone modification can have on the accessibility of the resulting chromatin. If the old histones were lost completely, then chromosome duplication would erase any "memory" of the previously modified nucleosomes. In contrast, if the old histones were retained on a single chromosome, that chromosome would have a distinct set of modifications relative to the other copy of the chromosome. In experiments that differentially labeled old and new histones, it was found that the old histones are present on both of the daughter chromosomes. Thus, as the replication fork passes, nucleosomes are broken down in to their component subassemblies. H2B dimers are released and enter the local pool, available for new nucleosome assembly. The distributive inheritance of old histones during chromosome duplication provides a mechanism for the propagation of the parental pattern of histone modification. By this mechanism, old modified histones will tend to rebind one of the daughter chromosomes at a position near their previous position on the parental chromosome. This localized inheritance of modified histones ensures that a subset of the modified histones is located in similar positions on each daughter chromosome. As the chromosome is replicated, histones that were associated with the parental chromosome are differently distributed. H4 tetramers are randomly transferred to one of the two daughter strands but do not enter in to the soluble pool of H3. H4 tetramers form the basis of the nucleosomes on the strand that does not inherit the parental tetramer. In contrast, H2A and H2B dimers are released in to the soluble pool and compete for H3. These tetramers will include all of the modifications added to the parental nucleosomes. H4 tetramers results in the daughter chromosomes receiving the same modifications as the parent. The ability of these modifications to recruit enzymes that perform the same modifications facilitates the propagation of the modification to the two daughter chromosomes. Such mechanisms are likely to play a critical role in the inheritance of chromatin states from one generation to another. Assembly of Nucleosomes Requires Histone "Chaperones" the assembly of nucleosomes is not a spontaneous process. For correct nucleosome assembly, it was necessary to raise salt concentrations to very high levels (. Although useful for assembling nucleosomes for in vitro studies (such as for the structural studies of the nucleosome described above), elevated salt concentrations are not involved in nucleosome assembly in vivo. After the replication fork has passed, chromatin assembly factors chaperone free H3. Chromosomes can be either circular or linear; however, each cell has a characteristic number and composition of chromosomes. In eukaryotic cells, centromeres play a critical role in the segregation of chromosomes, and telomeres help to protect and replicate the ends of linear chromosomes. Eukaryotic cells carefully separate the events that duplicate and segregate chromosomes as cell division proceeds. During mitosis, a highly specialized apparatus ensures that one copy of each duplicated chromosome is delivered to each daughter cell. A more compact form of chromatin, the 30-nm fiber, is readily formed by arrays of H1-bound nucleosomes. In addition, these complexes facilitate the Genome Structure, Chromatin, and the Nucleosome 255 exchange of H2A/H2B dimers. Nucleosome-remodeling complexes are recruited to particular regions of the genome to facilitate alterations in chromatin accessibility. A subset of nucleosomes is restricted to fixed sites in the genome and is said to be "positioned. Modification of the histone amino-terminal tails also alters the accessibility of chromatin. The types of modifications include acetylation and methylation of lysines, methylation of arginines, and phosphorylation of serines, threonines, and tyrosines. Acetylation of amino-terminal tails is frequently associated with regions of active gene expression and inhibits formation of the 30-nm fiber. Histone modifications alter the properties of the nucleosome itself, as well as acting as binding sites for proteins that influence the accessibility of the chromatin. In addition, these modifications recruit enzymes that perform the same modification, leading to similar modification of adjacent nucleosomes and facilitating the stable propagation of regions of modified nucleosomes/chromatin as the chromosomes are duplicated. Assembly involves the function of specialized histone chaperones that escort the H3. Thus, both chromosomes inherit modified histones that can then act as "seeds" for the similar modification of adjacent histones. Combinatorial complexity in chromatin structure and function: Revisiting the histone code. Where do these intergenic sequences come from and what are some of their functions Explain why each chromosome in a eukaryotic cell contains multiple origins of replication but includes one and only one centromere. For other reactions, you use nucleosomes that are methylated at lysine 36 of the histone protein H3. For a diploid human cell, state how many copies of each chromosome are present in each cell (or soon to be daughter cell). Which protein domain(s) recognize the acetylation of histone amino-terminal tails What type of protein domain do you predict allows the histone deacetylase to interact with the nucleosomes In our discussion of how templates act (Chapter 4), we emphasized that two identical surfaces will not attract each other.
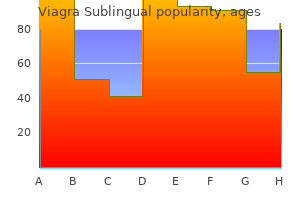
Purchase 100mg viagra sublingual free shipping
Cognitive theorists have argued that this mental representation may be best understood as an expectation of the unconditioned stimulus coffee causes erectile dysfunction generic viagra sublingual 100 mg fast delivery. In response to food, the dog not only salivates but also chews (if it is solid food) and swallows. Salivation becomes conditioned to the bell, but chewing and swallowing usually do not. Moreover, the bell comes to elicit not only salivation but also responses that do not usually occur in response to the food-in-mouth stimulus-such as tail wagging, food begging, and looking in the direction of the usual source of food (Jenkins et al. Rescorla summed up his cognitive view of classical conditioning as follows: "[Classical] conditioning is not a stupid process by which the organism willy-nilly forms associations between any two stimuli that happen to co-occur. Rather, the organism is best seen as an information seeker using logical and perceptual relations among events, along with its own preconceptions, to form a sophisticated representation of its world" (1988, p. By this, Rescorla does not mean that animals spend time consciously deliberating about these relationships. Rather, he means that animals have built-in neural mechanisms that automatically make the appropriate calculations. The sound of the can being attached to the opener permits her to predict the arrival of food. Her response is not identical to her response to food itself, but one of rapt attention. How do these observations support the idea that classical conditioning is a process of learning to predict the onset of the unconditioned stimulus Conditioning Depends on the Predictive Value of the Conditioned Stimulus Support for the expectancy theory of classical conditioning comes from research showing that conditioning occurs only, or at least mainly, when the new stimulus provides information that truly helps the animal predict the arrival of the unconditioned stimulus. Classical conditioning is most effective if the onset of the conditioned stimulus comes immediately before the unconditioned stimulus. Trying to achieve conditioning by placing the conditioned stimulus after the unconditioned stimulus is like trying to reduce traffic accidents by placing "Dangerous Curve" signs after the curves in the road rather than before them. The conditioned stimulus must signal heightened probability of occurrence of the unconditioned stimulus. Conditioning depends not just on the total number of pairings of the conditioned stimulus and unconditioned stimulus, but also on the number of times that either stimulus occurs without being paired with the other. As the number of pairings increases, conditioning is strengthened; but as the number of stimulus occurrences without pairing increases, conditioning is weakened (Rescorla, 1988; Rescorla & Wagner, 1972). This would be true no matter how many times, in our driving experience, we happened to come across such a sign right before an actual curve. If one conditioned stimulus already reliably precedes an unconditioned stimulus, a new stimulus, presented simultaneously with the original conditioned stimulus, generally does not become a conditioned stimulus. Even after many such pairings the new stimulus fails to elicit the conditioned response if it is presented alone. This failure of conditioning is called the blocking effect; the alreadyconditioned stimulus blocks conditioning to the new stimulus that has been paired with it (Kamin, 1969). In line with this interpretation, blocking does not occur if the new stimulus combined with the original stimulus is followed by an unconditioned stimulus that is larger in magnitude, or in some other way different from, the original unconditioned stimulus (Bradfield & McNally, 2008). In this case, the new stimulus does add new information, and the animal becomes conditioned to respond to it. Cognitive psychologists often emphasize that their use of terms such as expectation and prediction does not imply anything mystical. Some psychologists develop computer models to explain how such computations might be made. The three lower time lines illustrate patterns in which poor or no conditioning occurs even though the potential conditioned stimulus (blue) and unconditioned stimulus (red) are often paired. Strong conditioning occurs when the potential conditioned stimulus is a reliable and nonredundant predictor of the unconditioned stimulus. The potential conditioned stimulus occurs just before the unconditioned stimulus on most trials. The potential conditioned stimulus follows the unconditioned stimulus and is useless as a predictor. The potential conditioned stimulus often occurs before the unconditioned stimulus, but both stimuli also often occur at other times. One stimulus (green) is already established as a conditioned stimulus, so a new stimulus (blue) adds no further predictive value. A conditioned stimulus preceding a painful or startling event can elicit fear and bodily reactions that help brace the individual for that event; a conditioned stimulus preceding delivery of food can elicit hunger and bodily responses that help prepare the gastrointestinal tract for food; and a conditioned stimulus preceding an opportunity for sex can elicit high sex drive and bodily responses that help prepare the body for copulation. Watson was the first psychologist to show that the emotion of fear can be conditioned in human beings. Consistent with his behavioral perspective, Watson (1924) defined fear not as a feeling but as a set of observable responses: "a catching of the breath, a stiffening of the whole body, a turning away of the body from the source of stimulation, a running or crawling from it. In a classic demonstration, Watson and Rosalie Rayner (1920) conditioned an 11-month-old baby named Albert to fear laboratory rats. To condition the fear, the experimenters struck a steel bar with a hammer to produce a loud sound just after the rat was placed in front of Albert. After two such pairings Albert exhibited moderate fear in response to the rat alone, and after four more pairings he responded with strong fear to the rat alone. Thus, in the terminology of classical conditioning, the rat had become a conditioned stimulus for fear through being paired with a loud sound, which was an unconditioned stimulus for fear. The experiment most likely would not be approved by a modern ethics review committee and would not be allowed today. Moreover, Watson was far more interested in how to eliminate unwanted fears than in how to produce them. Fear, of course, is not the only emotional response that can be conditioned through Pavlovian procedures. After he was conditioned to respond fearfully to the rat, he also cried at the sight of other furry objects, including a rabbit, thereby exhibiting generalization (right). Some preferences seem to be instinctive-or, to use the term we introduced in Chapter 3, speciestypical-such as the human preferences for sweet flavors and social stimulation. Most, however, are learned; and this learning seems to occur primarily through classical conditioning. Evaluative conditioning refers to changes in the strength of liking or disliking of a stimulus as a result of being paired with another positive or negative stimulus. Levey and Irene Martin (1975), in which college students were shown picture postcards and asked to select their two favorite and their two least favorite. They were then paired with neutral cards (the conditioned stimuli)-some with the "liked" cards and others with the "disliked" cards-and the subjects were later asked to rate the cards again. This effect, using variants of this procedure for a wide range of stimuli, has been reported in hundreds of studies (De Houwer et al. It suggests that our likes and dislikes, which motivate much of our behavior, are governed to a significant degree by classical conditioning. Conditioning young humans to love beer the unconditioned stimulus is the happy, sexually suggestive scene. Conditioned Hunger A signal that reliably precedes food becomes a conditioned stimulus not just for salivation, but for a whole set of responses that help prepare the body for food and induce a state of hunger: the secretion of digestive juices in to the stomach, the secretion of certain hormones in to the bloodstream, and so on (Woods et al. The taste of a small morsel of food, the smell of food, a dinner bell, a clock indicating that it is dinnertime, or any other signal that reliably precedes a meal can rather quickly cause us to feel much hungrier than we were feeling just before the signal. Other animals, too, begin to act as if they are hungry, as indicated by their food-seeking behavior, when subjected to stimuli that have preceded food in the past (Pavlov, 1927/1960). In cases where a conditioned stimulus always precedes a specific kind of food, the conditioned hunger that occurs may be specific for that food. For example, in one series of experiments, rats that were made hungry through food deprivation underwent conditioning in which a tone always preceded the delivery of a certain kind of food. Later, when they were not food deprived, they would seek out and eat that same kind of food, but not other foods, when the tone sounded (Petrovich & Gallagher, 2007). The rats behaved as if the tone elicited a sudden craving for the specific food with which it had previously been paired when they were hungry. Purveyors of fast foods are capitalizing on classical conditioning when they plaster the environment with specific symbols that are associated over and over again with their specific foods. Conditioned Sexual Arousal A number of experiments have demonstrated that sexual arousal can be conditioned in nonhuman (Pfaus et al. Typically, a previously neutral stimulus is used as the conditioned stimulus (a pho to showing a jar of pennies was used in one experiment) and an erotic, sexually arousing film clip or sexually arousing vibration applied mechanically to the genitals is used as the unconditioned stimulus.
Thujone-containing Herbs (Mugwort). Viagra Sublingual.
- Dosing considerations for Mugwort.
- What is Mugwort?
- How does Mugwort work?
- Are there safety concerns?
- Stomach problems (colic, diarrhea, cramps, constipation, slow digestion, vomiting), epilepsy, irregular menstrual periods, low energy, anxiety, itching caused by scars, and other conditions.
Source: http://www.rxlist.com/script/main/art.asp?articlekey=96163
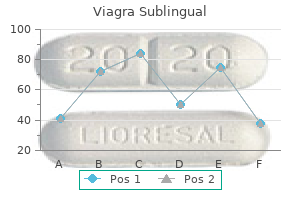
Order viagra sublingual 100mg without a prescription
Their goal was to isolate a small erectile dysfunction doctors in kansas city buy viagra sublingual discount, quickly reproducing animal that could be cultured in the lab and used to study the inheritance of quantitative traits, such as eye color. Among the menagerie of creatures that were captured, the fruit fly emerged as the animal of choice. Culturing was done in recycled milk bottles using an inexpensive concoction of yeast and agar. Drosophila Has a Rapid Life Cycle the salient features of the Drosophila life cycle are a very rapid period of embryogenesis, followed by three periods of larval growth prior to metamorphosis. Embryogenesis is completed within 24 hours after fertilization and culminates in the hatching of a first-instar larva. As we discussed in Chapter 21, the early periods of Drosophila embryonic development exhibit the most rapid nuclear cleavages known for any animal. A first-instar larva grows for 24 hours and then molts in to a larger, second-instar larva. The various stages of development of the fly, shown here, are described in the text. Disks are initially small and composed of fewer than 100 cells in the embryo but contain tens of thousands of cells in mature larvae. Imaginal disks differentiate in to their appropriate adult structures during metamorphosis (or pupation). The First Genome Maps Were Produced in Drosophila In 1910 the Morgan lab identified a spontaneous mutant male fly that had white eyes rather than the brilliant red seen for normal strains. These disks are initially formed as small groups of cells in the embryo but have grown to tens of thousands of cells in the mature larva. Sturtevant (a member of the Morgan lab), developed a simple mathematical algorithm for mapping the distances between linked genes based on recombination frequencies. The simplicity and power of this work had an enormous impact that fundamentally changed genetics and provided the first demonstration that genes are physically defined and ordered entities along the chromosomes. By the 1930s, extensive genetic maps were produced that identified the relative positions of numerous genes controlling a variety of physical characteristics of the adult, such as wing size and shape and eye color and shape. Muller, another scientist trained in the Morgan fly lab, provided the first evidence that environmental factors, such as ionizing radiation, can cause chromosome rearrangements and genetic mutations. The F1 progeny are heterozygous and contain one normal chromosome and one random mutation. In addition to its remarkable fecundity (a single female can produce thousands of eggs) and rapid life cycle, the fruit fly was found to possess several very useful features that guaranteed it a sustained and prominent role in experimental research. It contains only four chromosomes: two large autosomes, chromosomes 2 and 3, a smaller X chromosome (which determines sex), and a very small fourth chromosome. In the salivary gland, this process produces remarkable giant chromosomes composed of approximately 1000 copies of each chromatid. Bridges used these polytene chromosomes to determine a physical map of the Drosophila genome (the first produced for any organism). Bridges identified a total of approximately 5000 "bands" on the four chromosomes and established a correlation between many of these bands and the locations of genetic loci identified in the classical recombination maps. Endoreplication in the absence of cytokinesis generates enlarged chromosomes in some tissues of the fly, most notably the salivary glands where the giant chromosomes are composed of a thousand chromatids. It was possible, for the first time, to correlate the occurrence of genes for certain traits with specific physical segments of chromosomes. For example, white eye flies were correlated with deletions in the 3C region of the X chromosome. Balancer chromosomes (bottom panel) contain a series of inversions when compared with the original, parental chromosome (top panel). The left arm of the balancer chromosome has an internal inversion that reverses the order of genes a, b, and c in the original chromosome. Similarly, the arm on the right of the balancer chromosome has an inversion that reverses the order of genes d, e, and f. In addition, there might be an inversion centered around the centromere, in this case reversing the order of genes 1 and 2. The balancer chromosome thus has a significantly different order of genes when compared with the original. As a result, there is a suppression of recombination between the chromosomes in heterozygotes containing one copy of each. This type of analysis led to the conclusion that the white gene is located somewhere between polytene bands 3C2 and 3C3 on the X chromosome. A variety of additional genetic methods were created to establish the fruit fly as the premiere model organism for studies in animal inheritance. For example, balancer chromosomes were created that contain a series of inversions relative to the organization of the native chromosome. Critically, such balancers fail to undergo recombination with the native chromosome during meiosis. As a result, it is possible to maintain permanent cultures of fruit flies that contain recessive, lethal mutations. Consider a null mutation in the even-skipped (eve) gene, which we discussed in Chapter 21. Embryos that are homozygous for this mutation die and fail to produce viable larvae and adults. The null mutation can be maintained in a population that is heterozygous for a "normal" chromosome containing the null allele of eve and a balancer second chromosome, which contains a normal copy of the gene. Because the eve null allele is strictly recessive, these flies are completely viable. However, only heterozygotes are observed among adult progeny in successive generations. Embryos that contain two copies of the balancer chromosome die because some of the inversions produce recessive disruptions in critical genes. In addition, embryos that contain two copies of the normal chromosome die because they are homozgyous for the eve null mutation. Genetic Mosaics Permit the Analysis of Lethal Genes in Adult Flies Mosaics are animals that contain small patches of mutant tissue in a generally "normal" genetic background. Such small patches do not kill the individual because most of the tissues in the organism are normal. For example, small patches of engrailed/engrailed homozygous mutant tissue can be produced by inducing mitotic recombination in developing larvae using X-rays. When such patches are created in posterior regions of the developing wings, the resulting flies exhibit abnormal wings that have duplicated anterior structures in place of the normal posterior structures. The analysis of genetic mosaics provided the first evidence that Engrailed is required for subdividing the appendages and segments of flies in to anterior and posterior compartments. Individuals with two X chromosomes are females, whereas those with just one X are males. In all subsequent divisions, nuclei containing two X chromosomes give rise to daughter nuclei with two X chromosomes, whereas nuclei with just one X chromosome give rise to daughters containing a single X. As we discussed in Chapter 21, these nuclei undergo rapid cleavages without cell membranes and then migrate to the periphery of the egg. This migration is coherent and there is little or no intermixing of nuclei containing one X chromosome with Model Organisms 823 nuclei containing two X chromosomes. Thus, half the embryo is male and half is female, although the "line" separating the male and female tissues is random. Its exact position depends on the orientation of the two daughter nuclei after the first cleavage. The line sometimes bisects the adult in to a left half that is female and a right half that is male. If the wild-type X chromosome is lost at the first division, then the right half of the fly, the male half, has white eyes (the male half has only the mutant X chromosome), whereas the left half (the female side) has red eyes. It is composed of only approximately 150 Mb and contains fewer than 14,000 protein coding genes. As we discussed earlier, genetic mosaics are produced by mitotic recombination in somatic tissues.
Discount viagra sublingual american express
The van der Waals bonding energy between two atoms separated by the sum of their van der Waals radii increases with the size of the respective atoms erectile dysfunction treatment washington dc order 100mg viagra sublingual visa. For two average atoms, it is only 1 kcal/mol, which is just slightly more than the average thermal energy of molecules at room temperature (0. This means that van der Waals forces are an effective binding force at physiological temperatures only when several atoms in a given molecule are bound to several atoms in another molecule or another part of the same molecule. For several atoms to interact effectively, the molecular fit must be precise because the distance separating any two interacting atoms must not be much greater than the sum of their van der Waals radii. The strength of interaction rapidly approaches zero when this distance is only slightly exceeded. Thus, the strongest type of van der Waals contact arises when a molecule contains a cavity exactly complementary in shape to a protruding group of another molecule, as is the case with an antigen and its specific antibody. The bonding pattern of polar molecules is rarely dominated by van der Waals interactions because such molecules can acquire a lower energy state (lose more free energy) by forming other types of bonds. Sometimes, the hydrogen- 3-6 the arrangement of molecules in a layer of a crystal formed by the amino acid glycine. The structures, depicted as space filling (a) and as ribbons (b), show the complex between Fab D 1. The biologically most important hydrogen bonds involve hydrogen atoms covalently bound to oxygen atoms (O-H) or nitrogen atoms (N-H). In the absence of surrounding water molecules, bond energies range between 3 and 7 kcal/mol, the stronger bonds involving the greater charge differences between donor and acceptor atoms. Hydrogen bonds are thus weaker than covalent bonds, yet considerably stronger than van der Waals bonds. A hydrogen bond, therefore, will hold two atoms closer together than the sum of their van der Waals radii, but not so close together as a covalent bond would hold them. In the strongest hydrogen bonds, the hydrogen atom points directly at the acceptor atom. Hydrogen bonds are also much more specific than van der Waals bonds because they demand the existence of molecules with complementary donor hydrogen and acceptor groups. These charged groups are usually neutralized by nearby, oppositely charged groups. The electrostatic forces acting between the oppositely charged groups are called ionic bonds. When this happens in aqueous solution, the neutralizing cations and anions do not carry fixed positions because inorganic ions are usually surrounded byshells of water molecules and thus do not directly bind to oppositely charged groups. Thus, in water solutions, electrostatic bonds to surrounding inorganic cations or anions are usually not of primary importance in determining the molecular shapes of organic molecules. On the other hand, highly directional bonds result if the oppositely charged groups can form hydrogen bonds to each other. Because 3 these bonds are stronger than those that involve groups with less than a unit of charge, they are correspondingly shorter. A strong hydrogen bond can also form between a group with a unit charge and a group having less than a unit charge. This proximity is possible only when the molecular surfaces have complementary structures, so that a protruding group (or positive charge) on one surface is matched by a cavity (or negative charge) on another. The Importance of Weak and Strong Chemical Bonds 59 In cells, this requirement often means that some molecules hardly ever bond to other molecules of the same kind because such molecules do not have the properties of symmetry necessary for self-interaction. For example, some polar molecules contain donor hydrogen atoms and no suitable acceptor atoms, whereas other molecules can accept hydrogen bonds but have no hydrogen atoms to donate. On the other hand, there are many molecules with the necessary symmetry to permit strong self-interaction in cells. Instead, they exist as polar H-O-H molecules with both the hydrogen and oxygen atoms forming strong hydrogen bonds. In each water molecule, the oxygen atom can bind to two external hydrogen atoms, whereas each hydrogen atom can bind to one adjacent oxygen atom. In ice, the bonds to these neighbors are very rigid and the arrangement of molecules fixed. Above the melting temperature (08C), the energy of thermal motion is sufficient to break the hydrogen bonds and to allow the water molecules to change their nearest neighbors continually. Even in the liquid form, however, at any given instant most water molecules are bound by four strong hydrogen bonds. Weak Bonds between Molecules in Aqueous Solutions the average energy of a secondary, weak bond, although small compared with that of a covalent bond, is nonetheless strong enough compared with heat energy to ensure that most molecules in aqueous solution will form secondary bonds to other molecules. The energy gained by forming specific hydrogen bonds between water molecules favors the arrangement of the molecules in adjacent tetrahedrons. Although the rigidity of the arrangement depends on the temperature of the molecules, the pictured structure is nevertheless predominant in water as well as in ice. The specific structure of a solution at a given instant is markedly influenced by which solute molecules are present, not only because molecules have specific shapes, but also because molecules differ in which types of secondary bonds they can form. Thus, a molecule will tend to move until it is next to a molecule with which it can form the strongest possible bond. Because of the disruptive influence of heat, the specific configuration of a solution is constantly changing from one arrangement to another of approximately the same energy content. Equally important in biological systems is the fact that metabolism is continually transforming one molecule in to another and thus automatically changing the nature of the secondary bonds that can be formed. Organic Molecules That Tend to Form Hydrogen Bonds Are Water Soluble the energy of hydrogen bonds per atomic group is much greater than that of van der Waals contacts; thus, molecules will form hydrogen bonds in preference to van der Waals contacts. For example, if we try to mix water with a compound that cannot form hydrogen bonds, such as benzene, the water and benzene molecules rapidly separate from each other, the water molecules forming hydrogen bonds among themselves while the benzene molecules attach to one another by van der Waals bonds. It is therefore impossible to insert a non-hydrogen-bonding organic molecule in to water. Therefore, almost all of the molecules that cells acquire, either through food intake or through biosynthesis, are somewhat insoluble in water. Hydrophobic "Bonds" Stabilize Macromolecules the strong tendency of water to exclude nonpolar groups is frequently referred to as hydrophobic bonding. Some chemists like to call all of the bonds between nonpolar groups in a water solution hydrophobic bonds. But, in a sense, this term is a misnomer, for the phenomenon that it seeks to emphasize is the absence, not the presence, of bonds. For example, proteins are always constructed from L-amino acids, never from their mirror images, the D-amino acids (Box 3-1. Although the D- and L-amino acids have identical covalent bonds, their binding properties to asymmetric molecules are often very different. If an L-amino acid is able to attach to a specific enzyme, the D-amino acid is unable to bind. Most molecules in cells can make good "weak" bonds with only a small number of other molecules, partly because most molecules in biological systems exist in an aqueous environment. The formation of a bond in a cell therefore depends not only on whether two molecules bind well to each other, but also on whether bond formation is overall more favorable than the alternative bonds that can form with solvent water molecules. The nature of the chemical bond and the structure of molecules and crystals: An introduction to modern structural chemistry, 3rd ed. Hydrophobic bonds are important both in the stabilization of proteins and complexes of proteins with other molecules and in the partitioning of proteins in to membranes. They may account for as much as one-half of the total free energy of protein folding.
Discount viagra sublingual on line
Science is carried out by people who come to their research with certain expectations impotence yahoo answers buy generic viagra sublingual 100 mg online. Cues from observers led Hans to give responses that many misinterpreted as signs of vast knowledge. Types of Research Strategies In their quest to understand the mind and behavior of humans and other animals, psychologists employ a variety of research strategies. A useful way to categorize the various research strategies used by psychologists is to think of them as varying along the following three dimensions (Hendricks et al. The research design, of which there are three basic types-experiments, correlational studies, and descriptive studies. The setting in which the study is conducted, of which there are two basic types-field and laboratory. The data-collection method, of which there are two basic types-self-report and observation. Each of these dimensions can vary independently from the others, resulting in any possible combination of design, setting, and data-collection methods. Here we first describe the three types of research designs and then, more briefly, the other two dimensions. Research Designs the first dimension of a research strategy is the research design, which can be an experiment, a correlational study, or a descriptive study. Researchers design a study to test a hypothesis, choosing the design that best fits the conditions the researchers want to control. It might be a condition of the environment, such as temperature or amount of noise, or it might be a measure of behavior, such as a score on a test. In describing an experiment, the variable that is hypothesized to cause some effect on another variable is called the independent variable, and the variable that is hypothesized to be affected is called the dependent variable. The aim of any experiment is to learn whether and how the dependent variable is affected by (depends on) the independent variable. In psychology, dependent variables are usually measures of behavior, and independent variables are factors that are hypothesized to influence those measures. More specifically, an experiment can be defined as a procedure in which a researcher systematically manipulates (varies) one or more independent variables and looks for changes in one or more dependent variables while keeping all other variables constant. If all other variables are kept constant and only the independent variable is changed, then the experimenter can reasonably conclude that any change observed in the dependent variable is caused by the change in the independent variable. The people or animals that are studied in any research study are referred to as the subjects of the study. In other experiments, called betweengroups experiments (or sometimes, between-subjects experiments), there is a separate group of subjects for each different condition of the independent variable. Why were the subjects randomly assigned to the different treatments rather than allowed to choose their own treatment Example of a Within-Subject Experiment In most within-subject experiments, a number of subjects are tested in each condition of the independent variable, but within-subject experiments can also be conducted with just one subject. In each experiment, Pfungst tested Hans repeatedly, under varying conditions of the independent variable. In that experiment the independent variable was the presence or absence of blinders, and the dependent variable was the percentage of questions the horse answered correctly. Pfungst took care to keep other variables, such as the difficulty of the questions and the setting in which the questions were asked, constant across the two test conditions. This experiment is a within-subject experiment because it applied the different conditions of the independent variable (blinders) to the same subject (Hans). These researchers identified a group of patients suffering from major depression (defined in Chapter 16) and assigned them, by a deliberately random procedure, to different treatments. One group received both drug therapy and psychotherapy, a second received drug therapy alone, a third received psychotherapy alone, and a fourth received no scheduled treatment. In this experiment, the independent variable was the kind of treatment given, and the dependent variable was the 10 degree of depression after 16 weeks of treatment. This is a between-groups experiment because the manipulations of the 8 independent variable (that is, the different treatments used) 6 were applied to different groups of subjects. Notice that the researchers used a random method (a method 4 relying only on chance) to assign the subjects to the treatment groups. Random assignment is regularly used in between-group 2 experiments to ensure that the subjects are not assigned in a 0 way that could bias the results. If DiMascio and his colleagues Drug therapy and Drug therapy Psychotherapy No treatment had allowed the subjects to choose their own treatment group, psychotherapy those who were most likely to improve even without treatTreatment groups (independent variable) ment-maybe because they were more motivated to improve- might have disproportionately chosen one treatment condition over the others. With random therapy were the least depressed at assignment, any differences among the groups that do not stem from the differing the end of the 16-week treatment treatments must be the result of chance, and, as you will see later, researchers have period (according to the results of a statistical tools for taking chance in to account in analyzing their data. In contrast, convention in graphing experimental results, which is used throughout this book, subjects who received no treatment the figure depicts variation in the independent variable along the horizontal axis were the most depressed. As you can see in the figure, those in the drug-plus-psychotherapy group were the least depressed after the 16-week period, and those in the notreatment group were the most depressed. The results support the hypothesis that both drug therapy and psychotherapy help relieve depression and that the two treatments together have a greater effect than either alone. Correlational Studies Often in psychology we cannot conduct experiments to answer the questions because we cannot-for practical or ethical reasons-assign subjects to particular experimental conditions and control their experiences. Suppose, for example, that you are interested in the relationship between the disciplinary styles of parents and the psychological development of children. Perhaps you entertain the idea that frequent punishment is harmful, that it promotes aggressiveness or other unwanted characteristics. You might consider randomly assigning some families to a strict punishment condition and others to other conditions. A correlational study can be defined as a study in which the researcher does not manipulate any variable, but observes or measures two or more already existing variables to find relationships between them. Correlational studies can identify relationships between variables, which allow us to make predictions about one variable based on knowledge of another; but such studies do not tell us in any direct way whether change in one variable is the cause of change in another. She also rated the children on various aspects of behavior, such as cooperation and friendliness, through observations in their nursery schools. The main finding (discussed more fully in Chapter 12) was that children of authoritative parents scored better on the measures of behavior than did children of authoritarian or permissive parents. Thus, if parents would simply raise their children using an authoritative parenting style, their children would be more cooperative, friendly, and so forth. But because the study was not an experiment, we cannot justifiably come to that conclusion. The researcher did not control either variable, so we cannot be sure what was cause and what was effect. Some children may be better behaved than others for reasons quite separate from parental style, and parents with well-behaved children may simply glide in to an authoritative mode of parenting, while parents with more difficult children fall in to one of the other two approaches as a way of coping. For example, anything that makes families feel good about themselves (such as having good neighbors, good health, and an adequate income) might promote an authoritative style in parents and, quite independently, also lead children to behave well. Or maybe the causal variable has to do with the fact that children are genetically similar to their parents and therefore have similar personalities: the same genes that predispose parents to behave in a kind but firm manner may predispose children to behave well, and the same genes that predispose parents to be either highly punitive or neglectful may predispose children to misbehave. Although many correlational studies have found a relationship between viewing televised violence and displaying aggressive behavior, such studies cannot tell us whether television inspires the aggressive behavior or whether aggressive individuals are more likely than others to watch violent television programs. All too frequently, people-including even scientists who sometimes forget what they should know-use correlations to make unjustified claims of causal relationships on subjects including not only psychology, but health, economics, and more. However, that is a judgment based on logical thought about possible causal mechanisms or on evidence from other sources, not from the correlation itself (Rutter, 2007). That possibility seems far more plausible than any other possible explanation of the relationship between the two variables. Many correlational studies are analyzed in that way, and these studies are the ones most likely to be confused with experiments. In many other correlational studies, however, both variables are measured numerically and neither is used to assign subjects to groups. For example, a researcher might be interested in the correlation between the height of tenth-grade boys (measured in centimeters or inches) and their popularity (measured by counting the number of classmates who list the boy as a friend). In such cases, the data are assessed by a statistic called the correlation coefficient, which will be discussed in the section on statistical methods later in this chapter. Descriptive Studies Sometimes the aim of research is to describe the behavior of an individual or set of individuals without assessing relationships between different variables. As an example of one involving numbers, researchers might survey the members of a given community to determine the percentage who suffer from various mental disorders. As an example of a descriptive study not involving numbers, an animal behaviorist might observe the courtship behaviors of mallard ducks to describe in detail the sequence of movements that are involved. Some descriptive studies are narrow in focus, concentrating on one specific aspect of behavior, and others are broad, aiming to learn as much as possible about the habits of a particular group of people or species of animal.
Buy viagra sublingual once a day
Segments of the chain may be a helices or b strands; the rest have less regular conformations erectile dysfunction diabetes viagra order 100 mg viagra sublingual free shipping. We will outline ways to describe and classify possible tertiary structures in a subsequent section. Usually, the stabilities of the secondary and tertiary structures of a polypeptide chain depend on each other. Many proteins are composed of more than one polypeptide chain: quaternary structure refers to the way individual, folded chains associate with each other. We can distinguish cases in which there are a defined number of copies of a single type of polypeptide chain (generally called a "subunit" in this context, or a "protomer") and cases in which there are defined numbers of each of more than one type of subunit. Often, however, the tertiary or even secondary structures of the components of a protein oligomer. In other words, the individual subunits acquire secondary or tertiary structure only as they also acquire quaternary structure. One common example is the a-helical coiled-coil: two (or sometimes three or even four) polypeptide chains, either identical or different, adopt a-helical conformations and wrap very gently around each other. The individual chains are not, in general, stable as a helices on their own-if the oligomeric interaction is lost, the separated helices unravel in to disordered polypeptide chains. Note the ladder of hydrophobic side chains (mostly gray) at the interface between the two helices. If the polypeptide chain is too short, there are no conformations that bury enough hydrophobic groups to stabilize a folded structure. If the chain is too long, the complexity of the folding process is likely to generate errors. As a result of these restrictions, most stably folded conformations include between about 50 and 300 amino acid residues. Longer polypeptide chains generally fold in to discrete modules known as domains (see Box 6-2, Glossary of Terms); each domain generally falls within the 50- to 300-residue range just mentioned. The structures of individual domains of such a protein are similar to the structures of smaller, single-domain proteins. Each of the two or more domains of a folded polypeptide chain sometimes contains a continuous sequence of amino acid residues. Secondary structure: Elements of regular polypeptide-chain structure with main-chain hydrogen bonds satisfied. The secondary structures that occur frequently in proteins are the a helix and the parallel and antiparallel b sheets. Tertiary structure: the folded, three-dimensional conformation of a polypeptide chain. Quaternary structure: Multi-subunit organization of an oligomeric protein or protein assembly. Domain: A part of a polypeptide chain with a folded structure that does not depend for its stability on any of the remaining parts of the protein. Motif (sequence): A short amino acid sequence with characteristic properties, often those suitable for association with a specific kind of domain on another protein. Topology (or fold): the structure of most protein domains can be represented schematically by the connectivity in three dimensions of their constituent secondary-structural elements and the packing of those elements against each other. Jane Richardson introduced "ribbon diagrams," such as those in many of the figures in this chapter, as convenient ways to visualize the fold of a domain (see the caption to . Homologous domains (or proteins): Domains (or proteins) that derive from a common ancestor. They necessarily have the same fold, and they often (but not always) have recognizably similar amino acid sequences. Homology modeling: Modeling the structure of a domain based on that of a homologous domain. Ectodomain: the part of a single-pass membrane protein that lies on the exterior side of the cell membrane. Glycosylation: Addition of a chain, sometimes branched, of one or more sugars (glycans) to a protein side chain. The glycans can be N-linked (attached to the side-chain amide of asparagine) or O-linked (attached to the side-chain hydroxyl of serine or threonine). Denaturation: Unfolding a protein or a domain of a protein, either by elevated temperature or by agents such as urea, guanidinium hydrochloride, or strong detergent ("denaturants"). Chaperone: A protein that increases the probability of native folding of another protein, usually by preventing aggregation or by unfolding a misfolded polypeptide chain so that it can "try again" to fold correctly. Active site (or catalytic site): the site on an enzyme that binds the substrate(s), often in a configuration resembling the transition state of the reaction catalyzed. Allosteric regulation: Control of affinity or of the rate of an enzymatic reaction by a ligand that binds at a site distinct from that of the substrate(s). The mechanism of allosteric regulation often involves a change in quarternary structure-that is, a reorientation or repositioning of subunits with respect to each other. The intervening domain then looks like an insertion in to the domain that folds from the flanking segments. Basic Lessons from the Study of Protein Structures the large number of domain structures that have been determined experimentally allows us to draw the following conclusions. First, most hydrophobic side chains are, indeed, buried, and most polar side chains are exposed. Second, if a functional group that can donate or accept a hydrogen bond is buried, it almost always has a hydrogen-bonding partner. The reason for this property is easy to grasp, when we recall that were the polar group exposed on the domain surface, it would make a similar hydrogen bond with water (which can both donate and accept). If the hydrogen bond were missing in the folded conformation, a favorable energetic contribution would have been lost when water was stripped away from that group as the polypeptide chain folded. These hydrogen bonds are also satisfied in folded structures, in considerable part by formation of secondary structures. Both a helices and b sheets satisfy the main-chain hydrogen bonds of all of the residues within them. Polypeptide chains are shown here schematically as "ribbons"-a representation, introduced by Jane Richardson, that emphasizes the role of secondary structural elements in the folded conformation of a domain: a helices are curled ribbons; b strands are gently curved arrows, pointing toward the carboxyl terminus. Each of these domains is a b-sandwich with an immunoglobulin fold (see Box 6-3); the b strands of each domain are designated by letters in the order in which they follow in the polypeptide chain. Each domain has a single disulfide bond, shown in a stick representation with bonds to sulfur atoms in yellow. The colors run from dark blue at the amino terminus to green at the carboxyl terminus. The "rainbow" colors run from dark blue at the amino terminus to red at the carboxyl terminus. Moreover, one or more of those additional domains can fold from a polypeptide chain "inserted" between secondary structural elements of the principal domain. As a result, it is useful to classify the observed domain structures according to the kinds of secondary structures present within them. We observe that even a relatively short polypeptide chain could, in principle, have an astronomically large number of folded conformations. Only a restricted number of these appear in the large catalog of known 3D protein structures. These not only have a substantial proportion of their amino acid residues in a helices or b sheets (rather than in irregular loops, which would be much less likely to allow main-chain hydrogen bonding), but also have a relatively simple 3D folding pattern. Although there would be many ways for the polypeptide chain to pass from one of these eight or nine strands to the next, the observed pattern is one in which the chain makes either a sharp turn within one sheet, linking two adjacent strands, or passes across the top or bottom of the domain to the other sheet. One very important property of all known domain structures is that the chain does not form a knot-that is, if you imagined pulling on its ends, the whole thing would open in to a straight line. Classes of Protein Domains Classifications of protein domains allow simple, summary descriptions. The most important levels in the classification hierarchy are fold (also called topology) and homology. The fold class takes in to account not only the secondary structures, but also how the chain passes from one helix or strand to another.
Best viagra sublingual 100 mg
Researchers have found consistent cultural differences even in the ways that people perceive and remember aspects of their physical environment (discussed in Chapters 9 and 10) green tea causes erectile dysfunction purchase viagra sublingual overnight. The psychological specialty that explains mental experiences and behavior in terms of the culture in which the person developed is called cultural psychology. While social psychologists emphasize the immediate social influences that act on individuals, cultural psychologists strive to characterize entire cultures in terms of the typical ways that people within them feel, think, and act. For example, a cultural psychologist might contend that the frontier history of North America, in which individuals and families often had to struggle on their own with little established social support, helps explain why North Americans value independence and individuality so strongly. Concerning jealousy, a cultural psychologist would point to significant cultural differences in romantic and sexual mores. For example, some cultures are more tolerant of extramarital affairs than are others, and this difference affects the degree and quality of jealousy that is experienced. Some cultures have a strong double standard that condemns women far more harshly than men for sexual infidelity, and in those cultures violent revenge on the part of a jealous man toward his mate may be socially sanctioned (Bhugra, 1993; Vandello & Cohen, 2008). In other cultures, the same violence would dishonor the perpetrator and land him in jail. Four-year-olds behave differently from 2-year-olds, and middle-aged adults behave differently from adolescents. The psychological specialty that documents and describes the typical age differences that occur in the ways that people feel, think, and act is called developmental psychology. Developmental psychologists may describe the sequence of changes that occur, from infancy to adulthood, for any given type of behavior or mental capacity. At a superficial level, then, age itself can be an explanation: "She talks in such-andsuch a way because she is 3 years old, and that is how most 3-yearolds talk. Those processes include physical maturation of the body (including the brain), behavioral tendencies that are genetically timed to emerge at particular ages, the accumulated effects of many learning experiences, and new pressures and opportunities provided by the social environment or the cultural milieu as one gets older. At this deeper level, then, developmental psychology is an approach that brings together the other levels of analysis. Neural, physiological, genetic, evolutionary, learning, cognitive, social, and cultural explanations might all be brought to bear on the task of explaining behavioral changes that occur with age. Developmental psychologists are particularly interested in understanding how experiences at any given stage of development can influence behavior at later stages. A developmental analysis of jealousy might begin with a description of agerelated changes in jealousy that correspond with age-related changes in social relationships. Infants become jealous when their mother or other primary caregiver devotes extended attention to another baby (Hart, 2010). Children of middle-school age, especially girls, often become jealous when their same-sex "best friend" becomes best friends with someone else (Parker et al. These early forms of jealousy are similar in form and function to sexual jealousy, which typically emerges along with the first serious romantic attachment, in adolescence or young adulthood. Researchers have found evidence of continuity between early attachments to parents and friends and later attachments to romantic partners (discussed in Chapter 12). People who develop secure relationships with their parents and friends in childhood also tend, later on, to develop secure relationships with romantic partners, relatively untroubled by jealousy (Fraley, 2002; Main et al. Although true sexual jealousy is not seen until adolescence, children can become jealous over potential loss of friendship. Levels of Analysis Are Complementary these various levels of analysis provide different ways of asking questions about any psychological phenomenon, such as jealousy. However, these should not be viewed as alternative approaches to understanding, but rather as complementary approaches which, when combined, provide us with a more complete picture of important aspects of psychology. Although you may hear debates over the relative importance of genetic versus cultural influences on any behavior, even these most extreme levels of analysis should be viewed as complementary to one another, not as opposing poles of a philosophical argument. Genes are always expressed in a context, and culture constitutes an important component in that context. Wherever possible, throughout this book we will try to integrate findings from the various levels of analysis. To some degree, as indicated in the foregoing discussion, different psychological research specialties correspond to different levels of analysis. This is most true of the nine specialties already described: behavioral neuroscience, biopsychology, behavioral genetics, evolutionary psychology, learning psychology, cognitive psychology, social psychology, cultural psychology, and developmental psychology. For example, sensory psychology is the study of basic abilities to see, hear, touch, taste, and smell the environment; and perceptual psychology is the study of how people and animals make sense of, or interpret, the input they receive through their senses. Similarly, some psychologists identify their specialty as the psychology of motivation or the psychology of emotion. And, of course, many psychologists combine specialties; they may describe themselves as cognitive cultural psychologists, social neuroscientists, evolutionary developmental psychologists, and so forth. Two major specialties, which are closely related to each other, are devoted to the task of understanding individual differences among people. Personality psychologists and abnormal psychologists use various levels of analysis. Differences in the nervous system, in hormones, in genes, in learning experiences, in beliefs, in social pressures, or in cultural milieu may all contribute to an understanding of differences in personality and in susceptibility to particular mental disorders. Closely related to abnormal psychology is clinical psychology (discussed in Chapter 17), the specialty that is concerned with helping people who have mental disorders or less serious psychological problems. They offer psychotherapy or drug treatments, or both, to help people cope with or overcome their disorders or problems. Clinical psychologists who conduct research are usually interested in identifying or developing better treatment methods. They are simply convenient labels aimed at classifying, roughly, the different levels of analysis and topics of study that characterize the work of different research psychologists. Regardless of what they call themselves, good researchers often use several different levels of analysis in their research and may study a variety of topics that in some way relate to one another. Our main reason for listing and briefly describing some of the specialties here has been to give you an overview of the broad scope of psychological science. The Connections of Psychology to Other Scholarly Fields Another way to characterize psychology is to picture its place in the spectrum of disciplines that form the departments of a typical college of arts and sciences. One division is the natural sciences, including physics, chemistry, and biology, shown on the left side of the figure. The second division is the social sciences, including sociology, cultural anthropology, political science, and economics, shown on the right side of the figure. The third division is the humanities- including languages, philosophy, art, and music-shown in the lower part of the figure. Humans, unlike other animals, talk to one another, develop philosophies, and create art and music. On the natural science end, it is tied most directly to biology by way of behavioral neuroscience, behavioral genetics, and evolutionary psychology. On the social science end, it is tied most directly to sociology and cultural anthropology by way of social and cultural psychology. In addition to bridging the natural and social sciences, psychology ties the whole spectrum of sciences to the humanities through its interest in how people produce and understand languages, philosophies, art, and music. It should not be surprising that psychology has such meaningful connections to other disciplines. No wonder it is very often chosen as a second major, or as a minor, by students who are majoring in other fields. Many students reading this book are planning to major in psychology, but many others are majoring in other subjects. No matter what you have chosen as your major field, you are likely to find meaningful connections between that field and psychology. Psychology has more connections to other subjects taught in the university than does any other single discipline (Gray, 2008). Psychology as a Profession Psychology is not only an academic discipline but also a profession. The profession includes both academic psychologists, who are engaged in research and teaching, and practicing psychologists, who apply psychological knowledge and ideas in clinics, businesses, and other settings.
