

Buy atorlip-10 10 mg
This is particularly important for muscle fibers that do not extend the entire length of the muscle cholesterol levels what you need to know proven atorlip-10 10mg. The lateral transmission of force is also thought to stabilize the sarcolemma and to protect it from damage during contraction. The heavy chains are wound together in an -helical configuration to form a long rod-like segment, and the N-terminal portions of each heavy chain form a large globular head. The head region extends away from the thick filament toward the actin thin filament and is the portion of the molecule that can bind to actin. The other pair of light chains, called regulatory light chains, can be phosphorylated by Ca++/calmodulin-dependent myosin light chain protein kinase, which can influence the interaction of myosin with actin (see the section "Skeletal Muscle Types"). Myosin filaments form by a tail-to-tail association of myosin molecules, which results in a bipolar arrangement of the thick filament. The mechanisms controlling this highly organized structure of the myosin thick filament are not clear, although the cytoskeletal protein titin is thought to participate in the formation of a scaffold for organization and alignment of the thick filament in the sarcomere. Control of Skeletal Muscle Activity Motor Nerves and Motor Units Skeletal muscle is controlled by the central nervous system. The cell bodies of motor neurons are located in the ventral horn of the spinal cord. The motor axons exit via the ventral roots and reach the muscle through mixed peripheral nerves. The motor nerves branch in the muscle, and each branch innervates a single muscle fiber. The specialized cholinergic synapse that forms the neuromuscular junction and the neuromuscular transmission process that generates an action potential in the muscle fiber are described in Chapter 6. A motor unit consists of the motor nerve and all the muscle fibers innervated by the nerve. The motor unit is the functional contractile unit because all the muscle cells within a motor unit contract synchronously when the motor nerve fires. The size of motor units within a muscle varies, depending on the function of the muscle. Activation of varying numbers of motor units within a muscle is one way in which the tension developed by a muscle can be controlled (see "Recruitment" in the section "Modulation of the Force of Contraction"). The neuromuscular junction formed by the motor neuron is called an end plate (see Chapter 6 for details). Acetylcholine released from the motor neuron at the neuromuscular junction initiates an action potential in the muscle fiber that rapidly spreads along its length. The duration of the action potential in cardiac muscle, in contrast, is approximately 200 msec. The short duration of the skeletal muscle action potential allows very rapid contractions of the fiber and provides yet another mechanism by which the force of contraction can be increased. Increasing tension by repetitive stimulation of the muscle is called tetany (see the section "Modulation of the Force of Contraction"). A thick filament is formed by the polymerization of myosin molecules in a tail-to-tail configuration extending from the center of the sarcomere (A). This release causes intracellular [Ca++] to rise, which in turn promotes actin-myosin interaction and contraction. The time course for the increase in intracellular [Ca++] in relation to the action potential and development of force is shown in. The elevation in intracellular [Ca++] begins slightly after the action potential and peaks at approximately 20 msec. Although there is a gap (15 nm in width) between the T tubule and the terminal cisternae, proteins bridge this gap. On the basis of their appearance on electron micrographs, these bridging proteins are called feet. These feet are the Ca++ release channels in the membrane of the terminal cisternae that are responsible for the elevation in intracellular [Ca++] in response to the action potential. Calsequestrin is a low-affinity Ca++-binding protein that is present in the lumen of the terminal cisternae. It appears to bind triadin in a Ca++-dependent manner, which raises the possibility that it has a role more important than serving simply as a Ca++ buffer. Inhibition of Ca influx did not affect excitation-contraction coupling but did reduce maximal tetanic tension at high rates of electrical stimulation, which suggests that there may be some extrusion of intracellular Ca during tetany, which is compensated by Ca influx to maintain maximal tetanic tension. This pump is not unique to skeletal muscle; it is found in all cells in association with the endoplasmic reticulum. Phospholamban and sarcolipin are present in slow-twitch muscle, whereas myoregulin is present in both fast- and slow-twitch muscle. Once bound with Ca++, troponin C facilitates movement of the associated tropomyosin molecule toward the cleft of the actin filament. This movement of tropomyosin exposes myosin binding sites on the actin filament and allows a cross-bridge to form and thereby generate tension (see section "Cross-Bridge Cycling: Sarcomere Shortening"). These sites seem to be involved in controlling and enhancing the interaction between the troponin I and troponin T subunits. Although a given tropomyosin molecule extends over seven actin molecules, it is hypothesized that the strong binding of myosin to actin results in movement of an adjacent tropomyosin molecule, perhaps exposing myosin binding sites on as many as 14 actin molecules. This ability of one tropomyosin molecule to influence the movement of another may be a consequence of the close proximity of adjacent tropomyosin molecules. Such movement shortens the length of the sarcomere and thereby contracts the muscle fiber. The mechanism by which myosin produces force and shortens the sarcomere is thought to involve four basic steps that are collectively termed the cross-bridge cycle (labeled a to d in. Myosin next undergoes a conformational change termed "ratchet action" that pulls the actin filament toward the center of the sarcomere (state c). If intracellular [Ca++] is still elevated, myosin undergoes another cross-bridge cycle and produces further contraction of the muscle. The ratchet action of the cross-bridge is capable of moving the thin filament approximately 10 nm. As [Ca++] falls, Ca++ dissociates from troponin C, and the troponin-tropomyosin complex moves and blocks the myosin binding sites on the actin filament. As already noted, formation of the thick filaments involves the association of myosin molecules in a tail-totail configuration to produce a bipolar orientation. Such a bipolar orientation allows myosin to pull the actin filaments toward the center of the sarcomere during the cross-bridge cycle. The myosin molecules are also oriented in a helical array in the thick filament in such a way that cross-bridges extend toward each of the six thin filaments surrounding the thick filament. These myosin projections/cross-bridges can be seen on electron micrographs of skeletal muscle and appear to extend perpendicular from the thick filaments at rest. In the contracted state, the myosin cross-bridges slant toward the center of the sarcomere, which is consistent with the ratchet action of the myosin head. There is, however, uncertainty about how many myosin molecules contribute to the generation of force and whether both myosin heads in a given myosin molecule are involved. It has been calculated that there may be 600 myosin heads per thick filament, with a stoichiometry of 1 myosin head per 1. As a result of steric considerations, it is unlikely that all myosin heads can interact with actin, and calculations suggest that even during maximal force generation, only 20% to 40% of the myosin heads bind to actin. In isolated muscle preparations, maximum mechanical efficiency (65% efficiency) is obtained at a submaximal force of 30% maximal tension. In humans performing steady-state ergometer exercise, mechanical efficiencies range from 40% to 57%. The soleus muscle of the leg, in contrast, requires 90 msec to reach peak tension in response to an action potential, and then it relaxes slowly. The gastrocnemius muscle requires an intermediate time to reach peak tension (40 msec) because of the presence of both fasttwitch and slow-twitch muscle fibers in this muscle. Comparison of threebasic motorunit phenotypes in skeletalmuscle of extremitiesand trunk. Alterations in muscle mass and contractile phenotype in response to unloading models: role of transcriptional/ pretranslationalmechanisms. These two types of myosin isoforms have the same basic structure described previously, with two heavy chains and two pairs of light chains, although they differ in amino acid composition. It is very difficult to convert a slow-twitch muscle fiber into a fast-twitch fiber, although it can be accomplished by cross-innervation, which involves surgically interconnecting two motor neurons. Thus the motor innervation of the muscle fiber plays an important role in determining which type of myosin isoform is expressed in the muscle fiber. Further study showed that the intracellular Ca concentration in the muscle (secondary to differences in the activity pattern of the motor neuron) was an important determinant of whether the muscle fiber expressed the slow myosin isoform or the fast myosin isoform (see the section "Growth and Development").
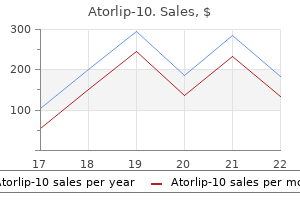
Order discount atorlip-10
Recently cholesterol chemical formula quality atorlip-10 10 mg, individuals have been found that have activating (gain-of-function) mutations in the V2 receptor gene. This new clinical entity has been termed nephrogenic syndrome of inappropriate antidiuresis. The sensation of thirst is satisfied by the act of drinking, even before sufficient water is absorbed from the gastrointestinal tract to correct the plasma osmolality. It is interesting to note that cold water is more effective in reducing the thirst sensation. Oropharyngeal and upper gastrointestinal receptors appear to be involved in this response. However, relief of the thirst sensation via these receptors is short lived, and thirst is only completely satisfied when the plasma osmolality or blood volume or pressure is corrected. However, most of the time fluid intake is dictated by cultural factors and social situations. In this situation, maintaining normal body fluid osmolality relies solely on the ability of the kidneys to excrete water. How the kidney accomplishes this is discussed in detail in the following sections of this chapter. Renal Mechanisms for Dilution and Concentration of Urine As already noted, water excretion is regulated separately from solute excretion. For this to occur the kidneys must be able to excrete urine that is either hypoosmotic or hyperosmotic with respect to body fluids. This ability to excrete urine of varying osmolality in turn requires that solute be separated from water at some point along the nephron. As discussed in Chapter 34, reabsorption of solute in the proximal tubule results in reabsorption of a proportional amount of water. Moreover, this proportionality between proximal tubule water and solute reabsorption occurs regardless of whether the kidneys excrete dilute or concentrated urine. Thus the proximal tubule reabsorbs a large portion of the filtered amount of solute and water, but it does not produce dilute or concentrated tubular fluid. The loop of Henle, in particular the thick ascending limb, is the major site where solute and water are separated. Thus excretion of both dilute and concentrated urine requires normal function of the loop of Henle. The nephron must simply reabsorb solute from the tubular fluid and not allow water reabsorption to also occur. Excretion of hyperosmotic urine is more complex and thus more difficult to understand. This mechanism is responsible for the polydipsia seen in response to the polyuria of both central and nephrogenic diabetes insipidus. Most individuals ingest water/beverages even in the absence of the thirst sensation. Normally the kidneys are able to excrete this excess water because they can excrete up to 18 L/day of urine. An example of how water intake can exceed the capacity of the kidneys to excrete water is long-distance running. A study of participants in the Boston Marathon found that 13% of the runners developed hyponatremia during the course of the race. Because over the course of the race they ingested (and generated through metabolism) more water than their kidneys were able to excrete or was lost by sweating, hyponatremia developed. In some racers the hyponatremia was severe enough to elicit the neurological symptoms described previously. For example, with maximally dilute urine (Uosm = 50 mOsm/kg H2O), the maximum urine output of 18 L/day will be achieved only if the solute excretion rate is 900 mmol/day: Uosm = Solute excretion Volume excreted 50 mOsm/kg H2O = 900 mmol/18 L If solute excretion is reduced, as commonly occurs in the elderly with reduced food intake, the maximum urine output will decrease. For example, if solute excretion is only 400 mmol/day, a maximum urine output (at Uosm = 50 mOsm/kg H2O) of only 8 L/day can be achieved. Thus individuals with reduced food intake have a reduced capacity to excrete water. Because water movement is passive, driven by an osmotic gradient, the kidney must generate a hyperosmotic compartment into which water is reabsorbed, without solute, osmotically from the tubular fluid. The hyperosmotic compartment in the kidney that serves this function is the interstitium of the renal medulla. Once established, this hyperosmotic compartment drives water reabsorption from the collecting duct and thereby concentrates urine. Establishment and maintenance of the hyperosmotic medullary interstitium has been a subject of study for more than 50 years. Despite this intense study, the most accepted model for how the medullary osmotic gradient is established, especially within the inner medulla, is incomplete and not consistent with more recent experimental findings regarding the transport properties of the nephron segments in this region of the kidney. With the caveat that the current model needs refinement, it is presented here because it embodies some fundamental concepts that underlie the process. In the current model the medullary interstitial osmotic gradient is established by a process termed countercurrent multiplication. This decreases the osmolality in the tubular fluid and raises the osmolality of the interstitium at this point. Thus at any point along the loop of Henle the fluid in the ascending limb has an osmolality less than fluid in the adjacent descending limb. Because of the countercurrent flow of tubular fluid in the descending (fluid flowing into the medulla) and ascending (fluid flow out of the medulla) limbs, this single effect could be multiplied, resulting in an osmotic gradient within the medullary interstitium, where the tip of the papilla has an osmolality of 1200 mOsm/kg H2O compared to 300 mOsm/kg H2O at the corticomedullary junction. Urea accumulates in the medullary interstitium (up to 600 mmol/L), which allows the kidneys to excrete urine with the same high urea concentration. Fluid entering the descending thin limb of the loop of Henle from the proximal tubule is isosmotic with respect to plasma. This reflects the essentially isosmotic nature of solute and water reabsorption in the proximal tubule (see Chapter 34). Most of this water is reabsorbed in the outer medulla, thereby limiting the amount of water added to the deepest part of the inner medullary interstitial space and thus preserving the hyperosmolality of this region of the medulla. In the inner medulla the terminal portion of the descending thin limb and all of the thin ascending limb is impermeable to water. This passive reabsorption of NaCl without concomitant water reabsorption begins the process of diluting the tubular fluid. The thick ascending limb of the loop of Henle is also impermeable to water and actively reabsorbs NaCl from the tubular fluid and thereby dilutes it further (see Chapter 34). Dilution occurs to such a degree that this segment is often referred to as the diluting segment of the kidney. The distal tubule and cortical portion of the collecting duct actively reabsorb NaCl. Under this condition, fluid leaving the cortical portion of the collecting duct is hypoosmotic with respect to plasma. The urine has an osmolality as low as approximately 50 mOsm/kg H2O and contains low concentrations of NaCl. An important point in understanding how a concentrated urine is produced is to recognize that while reabsorption of NaCl by the ascending thin and thick limbs of the loop of Henle dilutes the tubular fluid, the reabsorbed NaCl accumulates in the medullary interstitium and raises the osmolality of this compartment. Note also that during a water diuresis the osmolality of the medullary interstitium is reduced as a result of increased vasa recta blood flow and entry of some urea into the medullary collecting duct. This is thought to maintain the medullary interstitial gradient at a time when water is being added to this compartment from the medullary collecting duct, which would tend to dissipate the gradient. Because of NaCl reabsorption by the ascending limb of the loop of Henle, the fluid reaching the collecting duct is hypoosmotic with respect to the surrounding interstitial fluid. This diffusion of water out of the lumen of the collecting duct begins the process of urine concentration. The maximum osmolality the fluid in the distal tubule and cortical portion of the collecting duct can attain is approximately 290 mOsm/kg H2O. As the tubular fluid descends deeper into the medulla, water continues to be reabsorbed from the collecting duct, increasing the tubular fluid osmolality to 1200 mOsm/ kg H2O at the tip of the papilla. During antidiuresis, most of the water is reabsorbed in the distal tubule and cortical and outer medullary portions of the collecting duct.
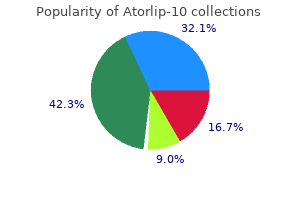
Cheap atorlip-10 uk
Thus when food reaches the stomach cholesterol ldl order line atorlip-10, protein digestion is initiated by generating protein hydrolysate, which further stimulates secretion of gastrin from the mucosa of the gastric antrum. In addition, gastric distention activates a vagovagal reflex that further stimulates gastric acid and pepsinogen secretion. Parietal cells express muscarinic receptors and are activated to secrete H+ in response to vagal efferent nerve activity. In addition, parasympathetic activation, via gastrin-releasing peptide from intrinsic neurons, releases gastrin from G cells located in the gastric glands in the gastric antrum. Gastrin enters the bloodstream and, via an endocrine mechanism, further stimulates the parietal cell to secrete H+. Thus gastrin and vagal efferent activity induce release of histamine, which potentiates the effects of both gastrin and acetylcholine on the parietal cell. Hence activation of parasympathetic (vagal) outflow to the stomach is very efficient at stimulating the parietal cell to secrete acid. In the gastric phase the presence of food in the stomach is detected and activates vagovagal reflexes to stimulate secretion. Food in the stomach results in distention and stretch, which are detected by afferent (or sensory) nerve endings in the gastric wall. These are the peripheral terminals of vagal afferent nerves that transmit information to the brainstem and thereby drive activity in vagal efferent fibers, a vagovagal reflex. In addition, digestion of proteins increases the concentration of oligopeptides and free amino acids in the lumen, which are detected by chemosensors in the gastric mucosa. Activation of vagal parasympathetic preganglionic outflow to the stomach acts in three ways to stimulate gastric acid secretion. There is direct neural innervation and activation of the parietal cell via release of acetylcholine (A) from enteric neurons, which acts on the parietal cell via muscarinic receptors. When the concentration of H+ in the lumen reaches a certain threshold (<pH 3), somatostatin is released from endocrine cells in the antral mucosa. Somatostatin has a paracrine action on neighboring G cells to decrease the release of gastrin and thereby decrease gastric acid secretion. The receptors on the parietal cell membrane for acetylcholine, gastrin, and histamine, as well as the intracellular second messengers by which these secretagogues act, are shown in. Thus much of the response to gastrin results from gastrinstimulated release of histamine. Acetylcholine binds to M3 muscarinic receptors and opens Ca++ channels in the apical plasma membrane. However, this is not required for full digestion of a meal; intestinal digestion is sufficient. Amylase is sensitive to pH and inactivated at low pH; however, some amylase is active even in the acidic gastric environment of the stomach because of substrate protection. Thus when carbohydrate occupies the active site of amylase, it protects the enzyme from degradation. Endocrine cells in the mucosa of the gastric antrum sense the presence of H+ and secrete somatostatin. This in turn acts on specific receptors on G cells to inhibit release of gastrin and thus bring about inhibition of gastric acid secretion. The terminals of the vagal preganglionic neurons innervate many enteric neurons and thus bring about changes in function as described in. The mixing patterns of gastric motility result in formation of an emulsion of lipids and gastric lipase, which attaches to the surface of lipid droplets in the emulsion and generates free fatty acids and monoglyceride from dietary triglyceride. However, the extent of hydrolysis of triglyceride is approximately 10%, and hydrolysis is not essential for normal digestion and absorption of dietary lipids. Moreover, as discussed in the next chapter, the products of lipolysis are not available for absorption in the stomach because of its low luminal pH. The mucus allows the pH of epithelial cells to be maintained at nearly neutral despite a luminal pH of about 2. Mucus also slows the diffusion of acid and pepsins to the epithelial cell surface. The motion of the gut wall governs the flow of the luminal contents along its length; the main patterns of motility are mixing (segmentation) and propulsion (peristalsis). In addition, smooth muscle activity in the stomach and colon subserves a storage function. Fusiform cells are packed together in bundles surrounded by a connective tissue sheath. Gap junctions functionally couple the smooth muscle cells so that contraction of bundles occurs synchronously. The frequency of slow waves is 3 to 5 per minute in the stomach and about 12 to 20 per minute in the small intestine; it decreases to 6 to 8 per minute in the colon. Their long processes form gap junctions with the longitudinal and circular smooth muscle cells; the gap junctions enable the slow waves to be conducted rapidly to both muscle layers. The slow wave will initiate a contraction in smooth muscle when it reaches a threshold amplitude. The amplitude of the slow wave is altered by release of neurotransmitters from enteric neurons. Gastric and duodenal erosions and ulcers occur as a result of an imbalance between the mechanisms that protect the mucosa and aggressive factors that can break it down. A healthy stomach/duodenum has ample natural protection against the destructive effects of H+. Infectious agents can also cause gastritis (inflammation of the gastric epithelium). The amplitude and, to a lesser extent, the frequency of the slow wave can be modulated by the activity of intrinsic and extrinsic nerves and by hormones and paracrine substances. If the depolarization of the slow wave exceeds the threshold, a train of action potentials may be triggered during the peak of the slow wave. The rising phase of the action potential is caused by flow of ions through channels that conduct both Ca++ and Na+ and are relatively slow to open. The Ca++ that enters the cell during the action potential helps initiate contraction. The extent of depolarization of the cells and the frequency of action potentials are enhanced by some hormones and paracrine agonists and by neurotransmitters from excitatory enteric nerve endings. Slow waves that are not accompanied by action potentials elicit little or no contraction of the smooth muscle cells. The greater the number of action potentials that occur at the peak of a slow wave, the more intense the contraction of the smooth muscle. Because smooth muscle cells contract rather slowly (about a 10th as fast as skeletal muscle cells), the individual contractions caused by each action potential in a train do not cause distinct twitches; rather they sum temporally to produce a smoothly increasing level of tension. Peristalsis occurs in the pharynx, esophagus, gastric antrum, and the small and large intestine. Segmental contractions produce narrow areas of contracted segments between relaxed segments. During spasm, maximal contractile activity occurs continuously in a dysregulated manner. In ileus, contractile activity is markedly decreased or absent; it often results from irritation of the peritoneum, such as occurs in surgery, peritonitis, and pancreatitis. In the gastric phase of the meal the pylorus is usually closed, and these antral contractions serve to mix the gastric contents and reduce the size of solid particles (grinding). However, eventually these antral contractions are also important in emptying the stomach of its contents. The pyloric sphincter is the gastroduodenal junction and is defined as an area of thickened circular muscle. Gastric Motility Functional Anatomy of the Stomach As discussed, the stomach is divided into two functional regions-proximal and distal, with sphincters at either end. By maintaining tone, reflux of contents from the stomach back into the esophagus is prevented. The proximal part of the stomach (the fundus together with the corpus or body) produces slow changes in tone compatible with its reservoir function.
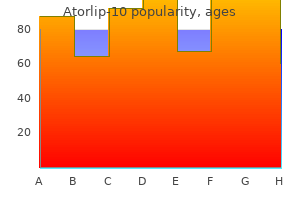
Buy discount atorlip-10 10 mg on-line
In the evolution of humans cholesterol lowering diet eggs order atorlip-10 10mg visa, these chemical senses apparently did not have the survival value of some of the other senses, but they contribute considerably to quality of life and food selection, and they are important stimulants of digestion. In other animals, the chemical senses have greater survival value, and their activation evokes a number of social behaviors, including mating, territoriality, and feeding. Distribution and Innervation of Taste Buds Taste buds are located on different types of taste papillae found on the tongue, palate, pharynx, and larynx. Types of taste papillae include fungiform and foliate papillae on the anterior and lateral aspects, respectively, of the tongue and circumvallate papillae on the base of the tongue. The sensitivity of different regions of the tongue for different taste qualities varies slightly because taste buds responding to each type of taste are widely distributed. Taste the stimuli that we commonly know as tastes are actually mixtures of five elementary taste qualities: salty, sweet, sour, bitter, and umami. The central processes of the afferent fibers enter the medulla, join the solitary tract, and synapse in the nucleus of the solitary tract. In some animals, including several rodent species, the secondorder taste neurons of the solitary nucleus project rostrally to the ipsilateral parabrachial nucleus. An unusual feature of the central gustatory pathway is that it is predominantly an uncrossed pathway (unlike the central somatosensory pathways, which are predominantly crossed). Olfaction the sense of smell is much better developed in some animals (macrosmatic animals) than in humans. The ability of dogs to track other animals on the basis of odor is legendary, as is the use of pheromones by insects to attract mates. From its opposite side, the cell gives off an unmyelinated axon that joins other olfactory nerve filaments and penetrates the base of the skull through openings in the cribriform plate of the ethmoid bone. These olfactory nerves synapse in the olfactory bulb, a portion of the cerebral hemisphere of the brain located at the base of the cranial cavity, just below the frontal lobe. Like taste cells, olfactory chemoreceptors have a short life span (about 60 days), and they are also continuously replaced. However, olfactory receptor cells are true neurons and, as such, are the only neurons that are continuously regenerated throughout life. The olfactory mucosa is exposed to odorant molecules by ventilatory air currents or from the oral cavity during feeding. The odorants are temporarily bound in mucus to an olfactory binding protein that is secreted by a gland in the nasal cavity. Olfactory coding resembles taste coding in that most natural odors are complex and consist of many molecules that excite a wide variety of olfactory chemoreceptors. Coding for a particular perceived odor depends on the responses of many olfactory chemoreceptors, and the strength of the odorant is represented by the overall amount of afferent neural activity. Central Pathways people avoid consuming spoiled food and detect dangerous situations. For example, an unpleasant odorant is added to odorless, colorless natural gas so that people can easily detect a leak. The olfactory mucosa also contains somatosensory receptors of the trigeminal nerve. When performing clinical tests of olfaction, clinicians must avoid activating these somatosensory receptors with thermal or noxious stimuli, such as the ammonia used in "smelling salts. It contains mitral cells, interneurons (granule cells; periglomerular cells), and distinct synaptic clusters (glomeruli; see. As the olfactory afferent fibers reach the olfactory bulb from the olfactory mucosa, they branch as they approach an olfactory glomerulus to synapse on the dendrites of mitral cells. Each glomerulus is the target of thousands of olfactory afferent fibers, but all the afferent fibers to a single glomerulus convey input from the one type of olfactory receptor. This is all the more remarkable because olfactory receptor cells are being regenerated continuously and new axons must therefore navigate their way to a correct glomerulus. Activity in a mitral cell depolarizes these inhibitory cells, and they in turn inhibit the original and adjacent glomeruli. Because each glomerulus is specialized by being the target of afferent fibers for a unique combination of odor qualities, this appears to be a way of enhancing stimulus contrast, much the way horizontal cells do in the retina. Olfactory Receptors the olfactory chemoreceptor cells are located in the olfactory mucosa, a specialized part of the nasopharynx. The c the conscious perception of flavor, particularly of foods, is the result of both olfactory and gustatory input based on directly inhaled odor, taste from the food as it is macerated in the mouth, and retronasal odor from the volatile molecules that are released by maceration and pass up into the nasal cavity from the pharynx. The axons of the mitral cells are shown exiting in the olfactory tract to the right. Within the olfactory tracts is a nucleus, called the anterior olfactory nucleus, that receives input from the olfactory bulb and projects to the contralateral olfactory bulb through the anterior commissure. As each olfactory tract approaches the base of the brain, it splits into the lateral and medial olfactory striae. Axons of the lateral olfactory stria synapse in the primary olfactory cortex, which includes the prepiriform cortex (and, in many animals, the piriform lobe). The medial olfactory stria includes projections to the amygdala, as well as to the basal forebrain. These structures are portions of, or directly connected to , the limbic system (see Chapter 10). Of note is that the olfactory pathway is the only sensory system that does not have an obligatory synaptic relay in the thalamus before signals reach the cortex. However, olfactory information does reach the mediodorsal nucleus of the thalamus, and it is then transmitted to the prefrontal and orbitofrontal cortex. In addition, via its intimate connections with limbic and, by extension, hypothalamic structures, it provides input to subconscious mechanisms related to emotions, memory, and sexual behavior. Light enters the eye through the cornea and lens and is focused on the retina, which lines the back of the eye. The cornea is the most powerful refractive surface, but the lens has a variable power that allows images of near objects to be focused on the retina. The iris regulates depth of field and the amount of illumination that enters the eye. Photoreceptors synapse on retinal bipolar cells, which in turn synapse on other interneurons and on ganglion cells. The optic disc, where the optic nerve leaves the retina, contains no photoreceptors and is therefore a blind spot. The portion of the retina with the highest degree of spatial resolution is the fovea and the surrounding macula. Rod photoreceptors have high sensitivity, do not discriminate among colors, and function best under low light levels. Color vision relies on the three types of cones that have different spectral sensitivities. Bipolar cells and many ganglion cells have concentric receptive fields with an on-center/off-surround or off-center/on-surround organization. Photoreceptor, bipolar, and horizontal cells respond to stimulation by modulating their membrane potential and their release of neurotransmitters, but ganglion cells respond by generating action potentials. The axons of ganglion cells in the temporal retina project to the brain ipsilaterally; those in the nasal retina cross in the optic chiasm. Because the lens inverts the image that falls on the retina, each side of the visual field is projected to the contralateral side of the brain for both eyes. Within the map, information from each eye maps to alternating adjacent points to create ocular dominance columns that extend vertically in the cortex. Striate cortical neurons outside of layer 4 respond best to bar or edge stimuli oriented in a particular way. Cells that "prefer" a particular stimulus orientation are grouped in orientation columns. Some in the inferotemporal cortex are influenced chiefly by P cells, and they function in form detection, color vision, and face discrimination. M cells influence regions of the middle temporal and parietal cortex, which function in motion detection and the control of eye movements. The pinna and auditory canal convey airborne sound waves to the tympanic membrane. The three small bones (ossicles) of the middle ear transmit the vibrations of the tympanic membrane to the oval window of the fluid-filled inner ear. Hearing is most sensitive at about 3000 Hz because of the dimensions of the auditory canal and the mechanics of the ossicles. The cochlea of the inner ear has three main compartments: the scala vestibuli, the scala tympani, and the intervening scala media (cochlear duct). The cochlear duct is bounded on one side by the basilar membrane, on which lies the organ of Corti, the sound transduction mechanism.

Buy atorlip-10 canada
There are approximately 280 million Hgb molecules per red blood cell cholesterol test validity discount atorlip-10 10 mg line, which provides an efficient mechanism to transport O2. Myoglobin, a protein similar in structure and function to Hgb, has only one subunit of the Hgb molecule. It aids in the transfer of O2 from blood to muscle cells and in the storage of O2, which is especially critical in O2-deprived conditions. Abnormalities of the Hgb molecule occur with mutations in the amino acid sequence. Factors that shift the oxyhemoglobin dissociation curve totheright(decreasedaffinityofHgbforO2)ortotheleft(increased affinity). Physiological Factors That Shift the Oxyhemoglobin Dissociation Curve the oxyhemoglobin dissociation curve can shift in numerous clinical conditions, either to the right or to the left. Oxyhemoglobin Dissociation Curve In the alveoli, the majority of O2 in plasma quickly diffuses into red blood cells and chemically binds to Hgb. This process is reversible, so that Hgb quickly gives up its O2 to tissue through passive diffusion (the concentration of O2 in Hgb decreases). The point on the curve at which Hgb is 50% saturated with O2 is called the P50, and it is 27 mm Hg in normal adults. When the affinity of Hgb for O2 increases, the curve is shifted to the left, which causes the P50 to decrease. Shifts to the right or left of the dissociation curve have little effect when they occur at O2 partial pressures within the normal range (80 to 100 mm Hg). However, at O2 partial pressures below 60 mm Hg (steep part of the curve), shifts in the oxyhemoglobin dissociation curve can dramatically influence O2 transport. This shifts the dissociation curve to the right, which has a beneficial effect by aiding in the release of O2 from Hgb for diffusion into tissues. An increase in body temperature, as occurs during exercise, shifts the oxyhemoglobin dissociation curve to the right and enables more O2 to be released to tissues, where it is needed because the demand increases. During cold weather, a decrease in body temperature, especially in the extremities (lips, fingers, toes, and ears), shifts the O2 dissociation curve to the left (higher Hgb affinity). In this instance, PaO2 may be normal, but release of O2 in these extremities is not facilitated. This causes the dissociation curve to shift to the left, which further prevents the unloading and delivery of O2 to tissues. Another gas, nitric oxide, has great affinity (200,000 times greater than O2) for Hgb, and it binds irreversibly to Hgb at the same site that O2 does. Thus nitric oxide is used therapeutically as an inhalant in patients with pulmonary hypertension to reduce pressure. Although nitric oxide poisoning is not common, the clinician should be cautious when administering nitric oxide therapy for long periods. Fetal Hemoglobin (HgbF) As discussed previously, HgbF has a greater affinity for O2 than does adult Hgb, and the oxyhemoglobin dissociation curve thus shifts to the left. Oxygen Saturation, Content, and Delivery Each Hgb molecule can bind up to four O2 atoms, and each gram of Hgb can bind up to 1. At 100% O2 capacity, the heme groups of the Hgb molecules are fully saturated with O2, and at 75% O2 capacity, three of the four heme groups are occupied. Binding of O2 to each heme group increases the affinity of the Hgb molecule to bind additional O2. Oxygen delivery from the lungs to tissues is dependent on several factors, including cardiac output, the Hgb content of blood, and the ability of the lung to oxygenate the blood. The actual O2 extracted from blood by the tissue is the difference between the arterial O2 content and the venous O2 content, multiplied by cardiac output. Under normal conditions, Hgb leaves the tissue 75% saturated with O2, and only about 25% is actually used by tissues. Hypothermia, relaxation of skeletal muscles, and an increase in cardiac output reduce O2 extraction. Conversely, a decrease in cardiac output, anemia, hyperthermia, and exercise increase O2 extraction. Thus anaerobic metabolism is stimulated and results in the increases in levels of lactate and H+ and the subsequent formation of lactic acid. In cases of severe hypoxia, the extremities, toes, and fingertips may appear blue-gray (cyanotic) because of lack of O2 and increased deoxyhemoglobin levels. There are four major types of tissue hypoxia (hypoxic hypoxia, circulatory hypoxia, anemic hypoxia, histotoxic hypoxia), discussed in detail in Chapter 23. Erythropoiesis Tissue oxygenation depends on the concentration of Hgb and thus on the number of red blood cells available in the circulation. Red blood cell production (erythropoiesis) in the bone marrow is controlled by the hormone erythropoietin, which is synthesized in the kidneys by cortical interstitial cells. Although Hgb levels are normally very stable, decreased O2 delivery, low Hgb concentration, and low PaO2 stimulate the secretion of erythropoietin. Chronic renal disease damages the cortical interstitial cells and thereby suppresses their ability to synthesize erythropoietin. This causes anemia, along with decreased Hgb because of the lack of erythropoietin. Erythropoietin replacement therapy using epoetin alfa (Epogen, Procrit) or darbepoetin alfa (Aranesp) effectively increases red blood cell production. This ratio is similar at the tissue to blood compartment, where it is referred to as the respiratory quotient. Regulation of Hydrogen Ion Concentration and Acid-Base Balance the H+ concentration (pH) has a dramatic effect on many metabolic processes within cells, and regulation of pH is essential for normal homeostasis. Also, pK is the negative logarithm of the overall dissociation constant for the reaction and has a logarithmic value of 6. Acidbase imbalances are also caused by metabolic disorders such as metabolic acidosis. Gases (nitrous oxide, ether, helium) that have a rapid rate of air-to-blood equilibration are perfusion limited. Tissue oxygenation is dependent on Hgb within red blood cells and subsequently the number (and production) of red blood cells, which is controlled by the hormone erythropoietin. Low O2 delivery, low Hgb concentration, and low PaO2 stimulate the secretion of erythropoietin in the kidneys. Tissue hypoxia occurs when insufficient amounts of O2 are supplied to the tissue to conduct normal levels of aerobic metabolism. A comparative meta-analysis of maximal aerobic metabolism of vertebrates: implications for respiratory and cardiovascular limits to gas exchange. Explain the role of central and peripheral chemoreceptors in regulating respiration. Compare and contrast the roles of chemoreceptors and pulmonary mechanoreceptors in regulating respiration. P eople breathe without thinking, and they can willingly modify their breathing pattern and even hold their breath. Control of ventilation includes the generation and regulation of rhythmic breathing by the respiratory center in the brainstem and its modification by the input of information from higher brain centers and from systemic receptors. Ventilatory Control: An Overview There are four major sites of ventilatory control: (1) the respiratory control center, (2) central chemoreceptors, (3) peripheral chemoreceptors, and (4) pulmonary mechanoreceptors/sensory nerves. The respiratory control center is located in the medulla oblongata of the brainstem and is composed of multiple nuclei that generate and modify the basic ventilatory rhythm. This center consists of two main parts: (1) a ventilatory pattern generator, which sets the rhythmic pattern, and (2) an integrator, which controls generation of the pattern, processes input from higher brain centers and chemoreceptors, and controls the rate and amplitude of the ventilatory pattern. Input to the integrator arises from higher brain centers, including the cerebral cortex, hypothalamus, limbic system including the amygdalae, and cerebellum. Central chemoreceptors are located in the central nervous system just below the ventrolateral surface of the medulla. Peripheral chemoreceptors are located on specialized cells in the aortic arch (aortic bodies) and at the bifurcation of the internal and external carotid arteries (carotid bodies) in the neck. Pulmonary mechanoreceptors and sensory nerve stimulation, in response to lung inflation or to stimulation by irritants or release of local mediators in the airways, modify the ventilatory pattern. The collective output of the respiratory control center to motor neurons located in the anterior horn of the spinal column controls the muscles of respiration, and this output determines the automatic rhythmic pattern of respiration. Motor neurons located in the cervical region of the spinal column control the activity of the diaphragm through the phrenic nerves, whereas other motor neurons located in the thoracic region of the spine control the intercostal muscles and the accessory muscles of respiration. In contrast to automatic respiration, voluntary respiration bypasses the respiratory control center in the medulla. The neural activity controlling voluntary respiration originates in the motor cortex, and signaling passes directly to motor neurons in the spine through the corticospinal tracts.
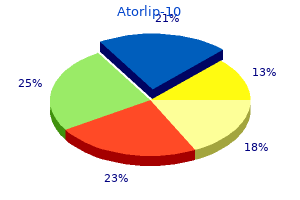
Order 10 mg atorlip-10 fast delivery
An increase in intraluminal pressure distends the vessels cholesterol levels calculator atorlip-10 10 mg sale, and a decrease reduces the caliber of the vessels as a consequence of elastic recoil of the vessel walls. At basal vascular tone, approximately a third of the blood volume of a tissue can be mobilized when the sympathetic nerves are stimulated at physiological frequencies. Extrinsic Control of Peripheral Blood Flow Sympathetic Neural Vasoconstriction Several regions in the cerebral medulla influence cardiovascular activity. Stimulation of the dorsal lateral medulla (pressor region) evokes vasoconstriction, cardiac acceleration, and enhanced myocardial contractility. Stimulation of cerebral centers caudal and ventromedial to the pressor region decreases arterial blood pressure. This depressor area exerts its effect by direct inhibition of spinal regions and by inhibition of the medullary pressor region. These areas are not true anatomical centers in which a discrete group of cells is discernible, but they constitute "physiological" centers. The effect of cholinergic fibers on total vascular resistance is small because only a small proportion of the resistance vessels of the body receive parasympathetic fibers. Thelate,slow, progressivedeclineinvolume(totherightofthearrow)iscausedby the movement of extravascular fluid into the capillaries and hence awayfromthetissue. A vasodilator polypeptide, bradykinin, formed locally from the action of an enzyme on a plasma protein substrate in the glandular lymphatic vessels, mediates this vasodilation. Bradykinin is formed in other exocrine glands, such as the lacrimal and sweat glands. Its presence in sweat may be partly responsible for the dilation of cutaneous blood vessels. Humoral Factors Epinephrine and norepinephrine exert a powerful effect on peripheral blood vessels. In skeletal muscle, low concentrations of epinephrine dilate resistance vessels (2-adrenergic effect), but high concentrations produce constriction (1adrenergic effect), as noted in Table 14. When stimulated, the adrenal gland can release epinephrine and norepinephrine into the systemic circulation. However, under physiological conditions, the effect of catecholamine release from the adrenal medulla is less important than norepinephrine release from sympathetic nerve endings. Local humoral substances have an important role in regulating vessel tone (Table 14. Therefore, at basal vascular tone, blood volume is close to the maximal blood volume of the tissue. More blood can be mobilized from the capacitance vessels in the skin than from those in the muscle. This disparity depends in part on the greater sensitivity of the skin vessels to sympathetic stimulation, but it also occurs because basal tone is lower in skin vessels than in muscle vessels. Therefore, in the absence of a neural influence, skin capacitance vessels contain more blood than do muscle capacitance vessels. For example, during physical exercise, activation of sympathetic nerve fibers constricts the peripheral veins and hence augments cardiac filling pressure. In arterial hypotension (as occurs in hemorrhage), the capacitance vessels constrict and thereby correct the decreased central venous pressure associated with blood loss. Vascular Reflexes Areas of the cerebral medulla that mediate sympathetic and vagal effects are under the influence of neural impulses that originate in the baroreceptors, chemoreceptors, hypothalamus, cerebral cortex, and skin. Arterial Baroreceptors Parasympathetic Neural Influence the efferent fibers of the cranial division of the parasympathetic nervous system innervate the blood vessels of the head and some of the viscera, whereas fibers of the sacral division innervate blood vessels of the genitalia, bladder, the baroreceptors (or pressoreceptors) are stretch receptors located in the carotid sinuses and in the aortic arch. Baroreceptor nerve terminals in the walls of the carotid sinus and aortic arch respond to the vascular stretch and deformation induced by changes in arterial blood pressure. The frequency of firing of these nerves is enhanced by an increase in arterial blood pressure and diminished by a reduction in arterial blood pressure. An increase in impulse frequency, as occurs with a rise in arterial pressure, inhibits the cerebral vasoconstrictor regions and results in peripheral vasodilation and lowering of arterial blood pressure. Bradycardia brought about by activation of the cardiac branches of the vagus nerves contributes to this lowering of blood pressure. Changes in carotid sinus pressure evoke greater changes in systemic arterial pressure and peripheral resistance than do equivalent changes in aortic arch pressure. The receptors in the carotid sinus walls respond more to pulsatile pressure than to constant pressure. At lower arterial pressure, these phasic changes are even more evident, but the overall discharge frequency is reduced. The blood pressure threshold for evoking sinus nerve impulses is approximately 50 mm Hg; maximal sustained firing is reached at approximately 200 mm Hg. Because the baroreceptors adapt, their response at any mean arterial pressure level is greater to a high pulse pressure than to a low pulse pressure. The increases in resistance that occur in response to reduced pressure in the carotid sinus vary from one peripheral vascular bed to another. The resistance changes elicited by altering carotid sinus pressure are greatest in the femoral vessels, less in the renal vessels, and least in the mesenteric and celiac vessels. Local application of norepinephrine or stimulation of sympathetic nerve fibers to the carotid sinuses enhances the sensitivity of its receptors in such a way that a given increase in intrasinus pressure produces a greater depressor response. Baroreceptor sensitivity decreases in hypertension because the carotid sinuses become stiffer as a result of the high intra-arterial pressure. Consequently, a given increase in carotid sinus pressure elicits a smaller decrease in systemic arterial pressure than it does at a normal level of blood pressure. Thus the set point of the baroreceptors is raised in hypertension in such a way that the threshold is increased and the pressure receptors are less sensitive to changes in transmural pressure. As would be expected, denervation of the carotid sinus can produce temporary and, in some instances, prolonged hypertension. The arterial baroreceptors play a key role in short-term adjustments in blood pressure in response to relatively abrupt changes in blood volume, cardiac output, or peripheral resistance (as in exercise). However, long-term control of blood pressure-over a period of days or weeks-is determined by the fluid balance of the individual: namely, the balance between fluid intake and fluid output. By far, the most important organ in the control of body fluid volume, and hence blood pressure, is the kidney (see also Chapter 35). Cardiopulmonary receptors are located in the atria, ventricles, and pulmonary vessels. Cardiopulmonary reflexes are tonically active and can alter peripheral resistance in response to changes in intracardiac, venous, or pulmonary vascular pressure. The atria contain two types of cardiopulmonary baroreceptors: those activated by the tension developed during atrial systole (type A receptors) and those activated by stretch of the atria during atrial diastole (type B receptors). Consequently, sympathetic activity is decreased to the kidney and increased to the sinus node. These changes in sympathetic activity increase renal blood flow, urine flow, and heart rate. Activation of the cardiopulmonary receptors can also initiate a reflex that lowers arterial blood pressure by inhibiting the vasoconstrictor center in the cerebral medulla. Stimulation of the cardiopulmonary receptors inhibits release of angiotensin, aldosterone, and vasopressin (antidiuretic hormone); interruption of the reflex pathway has the opposite effects. The reduction in blood volume (hypovolemia) enhances sympathetic vasoconstriction in the kidney and increases the secretion of renin, angiotensin, aldosterone, and vasopressin (see also Chapter 35). The renal vasoconstriction (primarily afferent arterioles) reduces glomerular filtration and increases release of renin from the kidney. The enhanced release of vasopressin decreases renal water excretion, and the release of aldosterone decreases renal NaCl excretion. These cardiac chemoreceptors are activated by ischemia of cardiac muscle, and they transmit the precordial pain (angina pectoris) associated with an inadequate blood supply to the myocardium. Hypothalamus Optimal function of the cardiovascular reflexes requires integrity of the pontine and hypothalamic structures. Furthermore, these structures are responsible for behavioral and emotional control of the cardiovascular system (see also Chapter 11). Stimulation of the anterior hypothalamus produces both a fall in blood pressure and bradycardia, whereas stimulation of the posterolateral region of the hypothalamus increases both blood pressure and the heart rate. The hypothalamus also contains a temperature-regulating center that affects blood vessels in the skin. Stimulation by the application of cold to the skin or by cooling of the blood perfusing the hypothalamus results in constriction of the skin vessels and heat conservation, whereas warm stimuli to the skin result in cutaneous vasodilation and enhanced heat loss.
Atorlip-10 10mg generic
Because the mechanism for protein reabsorption is "upstream" of the thick ascending limb cholesterol hdl ldl definition discount generic atorlip-10 uk. However, proteinuria in greater than trace amounts is often indicative of renal disease. Organic cations, including xenobiotics such as the antidiabetic agent metformin, the antiviral agent lamivudine, and the anticancer drug oxaliplatin, and many important monoamine neurotransmitters including dopamine, epinephrine, histamine, and norepinephrine are secreted by the proximal tubule. Uptake of organic cations is driven by the magnitude of the cell-negative potential difference across the basolateral membrane. These transport mechanisms are nonspecific, and several organic cations usually compete for secretion via a given transport pathway. Similar competition is observed for organic cation secretion by the proximal tubule, and elevated plasma levels of one transported cation species can inhibit secretion of the other competing cations. For example, the histamine H2 antagonist cimetidine used to treat gastric ulcers is secreted via organic cation transport mechanisms in the proximal tubule. If cimetidine is given to patients receiving procainamide (a drug used to treat cardiac arrhythmias), cimetidine reduces urinary excretion of procainamide (also an organic cation) by direct competition for a common secretory pathway. As a consequence, coadministration of cationic drugs competing for the same pathway can increase the plasma concentration of both drugs to levels much higher than those observed when the drugs are given alone. The positive voltage in the lumen plays a major role in driving the passive paracellular reabsorption of cations. Because the apical membrane is conductive primarily to K+, the apical membrane voltage is more negative than the basolateral membrane voltage, which is conductive to K+ and Cl-, thereby resulting in a lumen positive transepithelial potential. Reabsorption of water, but not NaCl, in the descending thin limb increases [NaCl] in the tubule fluid entering the ascending thin limb. As the NaCl-rich fluid moves toward the cortex, NaCl diffuses out of the tubule lumen across the ascending thin limb and into the medullary interstitial fluid, down a concentration gradient directed from the tubule fluid to the interstitium (see Chapter 35 for details). This transporter maintains a low intracellular [Na+], which provides a favorable chemical gradient for the movement of Na+ from tubular fluid into the cell. Using the potential energy released by the downhill movement of Na+ and Cl-, this symporter drives the uphill movement of K+ into the cell. These K+ channels allow the K+ transported into the cell via the 1Na+/1K+/2Cl- symporter to recycle back into tubule fluid. Because the [K+] in tubule fluid is relatively low, K+ recycling is required for continued operation of the 1Na+/1K+/2Cl- symporter. The operation of the Na+/H+ antiporter in the apical membrane results in cellular uptake of Na+ in exchange for H+. The voltage across the thick ascending limb is important for reabsorption of several cations. The tubular fluid is positively charged relative to blood because of the unique location of transport proteins in the apical and basolateral membranes. Two points are important: (1) increased NaCl transport by the thick ascending limb increases the magnitude of the positive voltage in the lumen, and (2) this voltage is an important driving force for reabsorption of several cations, including Na+, K+, Mg++, and Ca++, across the paracellular pathway. The importance of the paracellular pathway to solute reabsorption is underscored by the observation that inactivating mutations of the tight junction protein claudin-16 reduce reabsorption of Mg++ and Ca++ by the ascending thick limb, even in the presence of a lumen positive transepithelial voltage. In summary, NaCl reabsorption across the thick ascending limb occurs via transcellular and paracellular pathways. Because the thick ascending limb does not reabsorb water, owing to a lack of water channels. The initial segment of the distal tubule (early distal tubule) reabsorbs Na+, Cl-, and Ca++ and is impermeable to water. A number of proteins have now been identified as components of the tight junction, including proteins that span the membrane of one cell and link to the extracellular portion of the same molecule in the adjacent cell. Of these junctional proteins, claudins appear to be major determinants of the permeability characteristics of tight junctions. Claudin-2 is permeable to water and may be responsible for paracellular water reabsorption across the proximal tubule. Claudin-4 has been shown in cultured kidney cells to control the permeability of the tight junction to Na+, whereas claudin-15 determines whether a tight junction is permeable to cations or anions. Thus the permeability characteristics of the tight junctions in different nephron segments are determined at least in part by the specific claudins expressed by the cells in that segment. Thus dilution of tubular fluid begins in the thick ascending limb and continues in the early segment of the distal tubule. The last segment of the distal tubule (late distal tubule) and the collecting duct are composed of three cell types: principal cells and two types of intercalated cells. Reabsorption of Na+ generates a negative luminal voltage across the late distal tubule and collecting duct, which provides the driving force for paracellular reabsorption of Cl-. A variable amount of water is reabsorbed across principal cells in the late distal tubule and collecting duct. Although the negative potential inside these cells favors intracellular K+ retention, the electrochemical gradient across the apical membrane promotes secretion of K+ from the cell into tubular fluid (see Chapter 36). As a consequence these distal nephron segments possess the ability to both secrete and reabsorb K+ via independently regulated mechanisms, which contrasts with the general tendency to reabsorb Na+ along most nephron segments. However, other hormones (including dopamine and adrenomedullin), Starling forces, and the phenomenon of glomerulotubular balance also influence NaCl reabsorption. Taken together, these actions increase Na+ uptake across the apical cell membrane and facilitate Na+ exit from the cell interior into blood. The increase in reabsorption of Na+ generates a negative transepithelial luminal voltage across the late distal tubule and the collecting duct. This negative voltage in the lumen provides the electrochemical driving force for reabsorption of Cl- across the tight junctions. Aldosterone secretion is decreased by hypokalemia and natriuretic peptides (discussed in more detail next). Through its stimulation of NaCl reabsorption in the collecting duct, aldosterone also indirectly increases water reabsorption by this nephron segment. As noted earlier, aldosterone stimulates both NaCl reabsorption and K+ secretion by the collecting duct. By contrast, during hyperkalemia, K+ excretion by the kidneys is increased to normalize plasma [K+], albeit without an accompanying change in NaCl excretion. This phenomenon-the apparent independent effects of aldosterone on urinary Na+ and K+ excretion-is called the aldosterone paradox. The corresponding integrated physiological response to hyperkalemia is depicted in. Because the early distal tubule is not directly responsive to aldosterone, this hormone does not stimulate NaCl reabsorption in this segment. Urodilatin is secreted by the distal tubule and collecting duct and is not present in the systemic circulation; thus urodilatin influences only the function of the kidneys. It inhibits NaCl and water reabsorption across the medullary portion of the collecting duct. Uroguanylin and guanylin are produced by neuroendocrine cells in the intestine in response to oral ingestion of NaCl. Studies in Sgk1 knockout mice reveal that this kinase is required for animals to survive severe NaCl restriction and K+ loading. NaCl restriction and K+ loading enhance plasma [aldosterone], which rapidly (in minutes) increases Sgk1 protein expression and phosphorylation. These mutations increase the number of Na+ channels in the apical cell membrane of principal cells and thereby the amount of Na+ reabsorbed. The cause of the autosomal dominant form is an inactivating mutation in the mineralocorticoid receptor. First, NaCl and water reabsorption by the nephron (especially the proximal tubule) falls. Second, aldosterone secretion decreases, thus reducing NaCl reabsorption in the thick ascending limb, distal tubule, and collecting duct. Third, because angiotensin is a potent vasoconstrictor, a reduction in its concentration permits the systemic arterioles to dilate and thereby lower arterial blood pressure. The involvement of these gut-derived hormones helps explain why the natriuretic response of the kidneys to an oral NaCl load is more pronounced than when delivered intravenously. Catecholamines released from the sympathetic nerves (norepinephrine) and the adrenal medulla (epinephrine) stimulate reabsorption of NaCl and water by the proximal tubule, thick ascending limb of the loop of Henle, distal tubule, and collecting duct.
